蛋白質純化
其他連結
![]()
親和層析法
|
|
||||
|
|
蛋白質純化 |
其他連結 |
||
|
|
||||
|
親和層析法 |
||||
|
|
|
|
|
|
|
|
蛋白質純化 |
|
|
|
親和層析法 |
|||
|
.二級鍵 |
|||
|
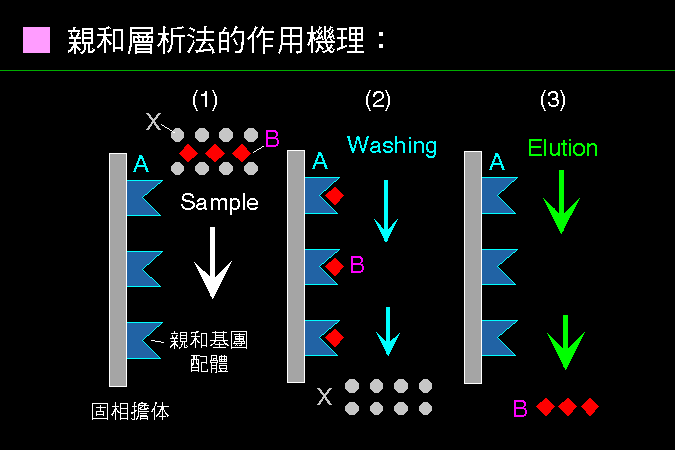
■ 親和層析法的作用機理 |
||
|
圖 例 |
||
|
|
||
|
親和層析法把目標蛋白質專一性地吸附住,洗去雜質後再溶離出來。 |
||
|
親和層析法有點像在釣魚,魚是我們所要的分子 (B) 雜夾在一堆蛋白質當中,魚餌是與 B 具有專一性的分子 (A, 上圖 1);A 與 B 的專一性很高,只會在樣本中與 B 結合,而排除其他雜質 (上圖 2);若把結合在吸著劑上的 B 溶離下來,就可以得到均質的 B (上圖 3)。 |
||
| P4A1 | ||
|
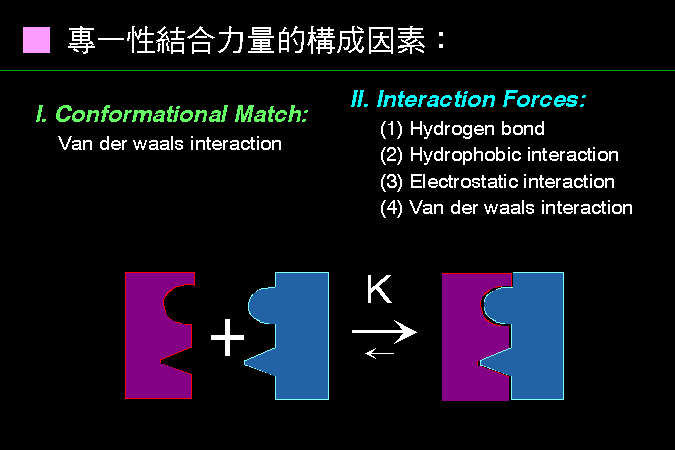
■ 專一性結合力量的構成因素 |
||
|
圖 例 |
||
|
|
||
|
有兩大類力量,構成分子之間的專一性吸引力。 |
||
|
一是兩個分子之間,其構形有如積木般的互補,會因為凡得瓦爾力的關係,產生專一性吸引力 (見下圖)。另一則為兩分子之間,因為某些基團間產生了二級鍵,所造成的吸引力。通常兩個分子間的專一性吸引力的形成,這兩大類力量都會有貢獻。 親和性結合力量的大小,可以用解離常數來表示,一般解離常數至少在 10-6 以上才算有足夠的親和力 (也就是說一百萬對結合分子中,只有一對會解離開來)。 |
||
| P4A2 | ||
|
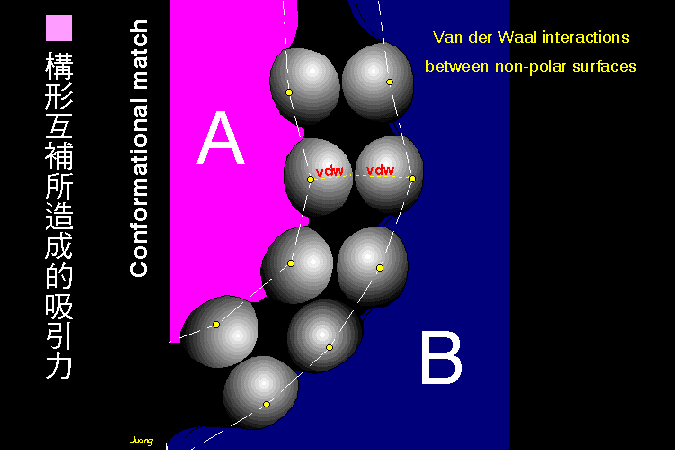
■ 構形互補所造成的吸引力 |
||
|
圖 例 |
||
|
|
||
|
凡得瓦爾力 是造成兩個互補平面之間產生專一性吸引力的主因。 |
||
|
凡得瓦爾力 是分子間微弱且短暫的吸引力,多發生在非極性的分子之間;雖然極性分子之間也會有凡得瓦爾吸引力,但其重要性則不若其它二級鍵。 繞在原子最外圍的電子依一定的軌道運轉,而此電子在軌道球面上的分佈通常是均勻的。當兩個原子漸漸接近時,這兩個原子上的電子會相互影響其分佈情形;若其中一個原子的電子分佈非常短暫地發生不均勻現象,則會導致短暫的偶極性產生 (d+.d-),而這短暫的偶極性馬上誘導另一個原子上的電子,也產生電子分佈不均的現象,但偶極剛好相反 (d-.d+)。 這兩個相反的偶極,便可以產生極微弱且短暫的吸引力,即稱為凡得瓦爾力。 凡得瓦爾力的存在太短暫,因此很容易就散掉;為了達致最大的凡得瓦爾力,兩個原子核之間,要保持在一最適距離;太近則因電子相斥而減弱,太遠則電子間的吸引力又不夠。這個最適距離,即為該原子的凡得瓦爾半徑。 又因為凡得瓦爾力實在很弱,因此要集合數十個凡得瓦爾力才能發揮作用。兩個巨分子間的接面,若其上的對應原子們,每一對都能保持在凡得瓦爾半徑上,則此二巨分子便可產生相當強的親和力量;這只有當兩個巨分子間的接觸面,剛好成為積木般的互補形狀時,才能達成。這種巨分子間的親和力,造就了非常多的細胞生理特性與功能,極為重要。 |
||
| P4A3 | ||
|
■ 二級鍵︰次於共價鍵的結合能力 |
||
|
圖 例 |
||
|
|
||
|
二級鍵是構成生物分子間各種功能或活性最重要的鍵結力量。 |
||
|
同一分子內的原子間是以共價鍵連接,而分子與分子之間,則是靠一些次於共價鍵的微弱二級鍵,作為聯繫兩個分子的力量。二級鍵的形成,事實上都與電子有關。 離子鍵是正電基團與負電基團間的吸引力;氫鍵是氫的電子被奪而露出質子,與附近的氧或氮原子上的電子產生吸引力;凡得瓦爾力是兩原子間,電子分佈不均所形成的微弱正負引力。 二級鍵的能量都很弱,約只有共價鍵的百分之一到千分之一;因此,兩分子間若要以二級鍵形成有效的鍵結,就要增加其總鍵數。二級鍵的好處是,不用費太大的能量就可打斷,因此適合可逆性的作用模式。 想像一個柔軟又可運動的人體,若全身都是由共價鍵所組成,則此人體每次動作,一定要先打斷一堆共價鍵,然後在動作完成後,再形成另一堆共價鍵。這種過程勢必耗費極大能量,不切實際。是故細胞內許多大小分子之間的結合,多不是以共價鍵為之,而是使用較弱但數目較多的二級鍵。 |
||
| P4A4 | ||
|
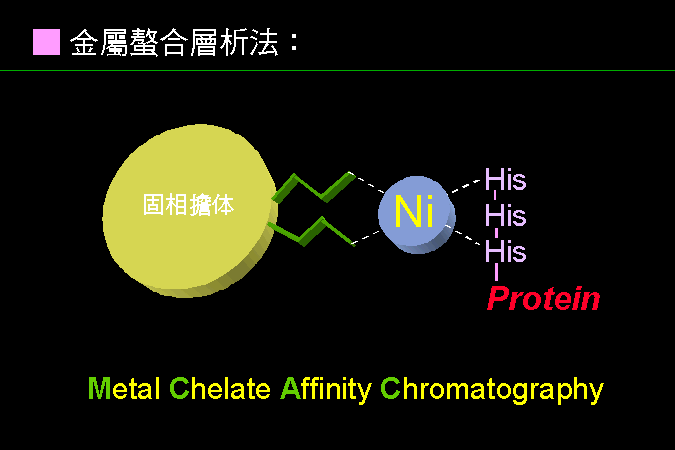
■ 金屬螯合層析法 |
||
|
圖 例 |
||
|
|
||
|
會與金屬結合的蛋白質,可利用金屬做為專一性螯合基團。 |
||
|
若在固相擔體接上金屬,則此金屬可被某些蛋白質所螯合,當洗去雜質後,即可純化出目標蛋白質。這類蛋白質分子中,多需要金屬做為 cofactor;也有特地在分子選殖時,加上一段可以與金屬結合的片段。 例如上例中,若在蛋白質端點接上一段含有六個 His 的片段,則此蛋白質可結合到接有金屬鎳的吸著劑。BCT 課程中的 GUS 已經接有這種 (His)6 的 tag,可以用金屬螯合法快速純化之。 |
||
| P4A5 | ||
▼ 下一小節︰ 純化策略
|
構 造 . 純 化 . 分 析 |
|
|
|
Protein/Purification/P4 |
|
本網頁最近修訂日期: 2001/06/30 |