蛋白質分析
其他連結
![]()
免疫轉印法
|
|
||||
|
|
蛋白質分析 |
其他連結 |
||
|
|
||||
|
免疫轉印法 |
||||
|
|
|
|
|
|
|
|
蛋白質分析 |
|
|
|
免疫轉印法 |
|||
|
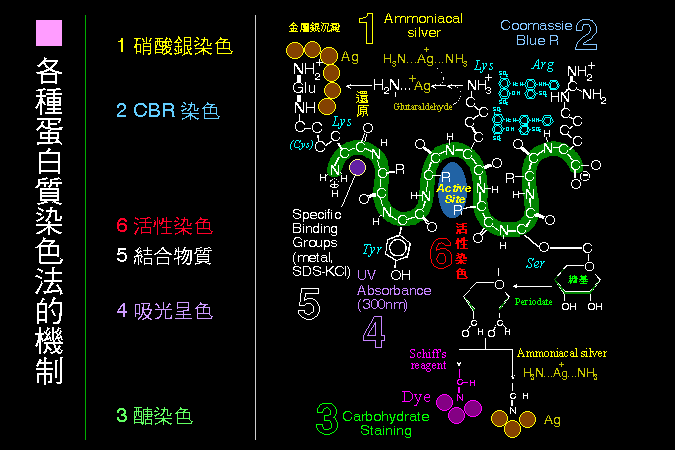
■ 各種蛋白質染色法的原理 |
||
|
圖 例 |
||
|
|
||
|
蛋白質染色也是利用到蛋白質分子上的各種基團。 |
||
|
與 蛋白質定量法 的原理大致相同,跑完電泳後,蛋白質的各種染色方法,也是利用蛋白質分子上的種種特性,尤其是各種胺基酸的基團。以下是最常用的三類膠片染色方法,但若要進行蛋白質轉印,就不能先進行這些染色法,因為染色後蛋白質都被固定在膠片上。 最方便而常用的染色,是使用 Coomassie Blue R 與蛋白質上面的 (1) 非極性基團 (2) Arg 或 Lys 等鹼性基團結合,因而使得蛋白質染上藍色。 硝酸銀染色法 的靈敏度比 Coomassie Blue 高至百倍,但步驟較為繁複,而且有些蛋白質會有負染色的情形;硝酸銀染色可能是銀銨錯離子與蛋白質上面的 Lys, Arg 等鹼性基團結合,然後被還原成金屬銀而呈色。 蛋白質上面的糖分子,可被過碘酸 (periodic acid) 氧化產生醛基,後者可與 Schiff's reagent 結合生成洋紅色的堆積物,因而染成洋紅色,此稱為 PAS 糖染色。 上述醛基也可以與硝酸銀染色的銀銨錯離子結合,因而染上金屬銀,因此也可以把硝酸銀染色用來作為糖染色。 |
||
| A4A1 | ||
|
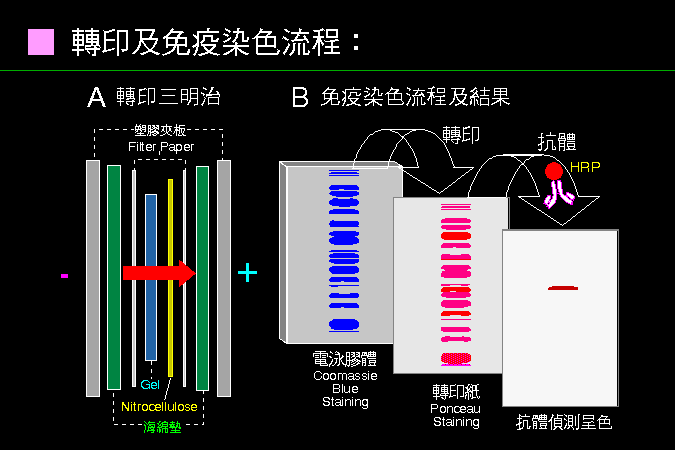
■ 轉印及免疫染色流程 |
||
|
圖 例 |
||
|
|
||
|
蛋白質轉印原理很簡單,但後續的染色操作及結果解讀要很小心。 |
||
|
通常蛋白質轉印不很困難,只要照上面把膠片-轉印紙三明治包好,通電後即可轉印成功。但再來使用抗體的免疫染色,就要小心各個操作步驟,以免染出來一片空白,或更慘的是,所有色帶全部都被染出來。 |
||
| A4A2 | ||
|
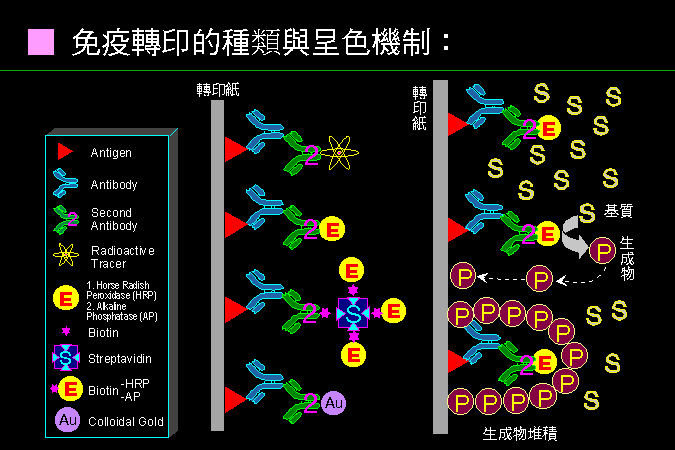
■ 免疫轉印的種類與呈色機制 |
||
|
圖 例 |
||
|
|
||
|
免疫染色要靠專一性高的抗體,以及靈敏的二次抗體-酵素連結體。 |
||
|
免疫染色是利用抗體來偵測轉印紙上面的抗原蛋白質,其原理也相當單純,但有很多應用上或技術上的問題。上圖列出目前較常用的免疫染色方法,都是源自最早的放射線免疫呈色,但現在都改用酵素,並且使所產生的生成物為水不溶性,以便把呈色堆積在轉印紙上。 為了提高靈敏度,有許多方法可用;例如酵素改用較靈敏的 alkaline phosphatase,或更採用會產生螢光產物的基質,靈敏度直逼放射性標示。更可使用 avidine (上圖 S) 及 biotin (上圖洋紅色六角星) 的放大系統,把單一個抗體的結合,連接到更多酵素標示,可提高靈敏度至百倍。 若使用金的膠體作為二次抗體的標示,則也可以用在電子顯微鏡的組織切片染色上,追蹤抗原分子的細胞位置;甚至可使用不同大小的金膠體,分別標示在兩種抗體,追蹤不同的抗原蛋白質。 |
||
| A4A3 | ||
|
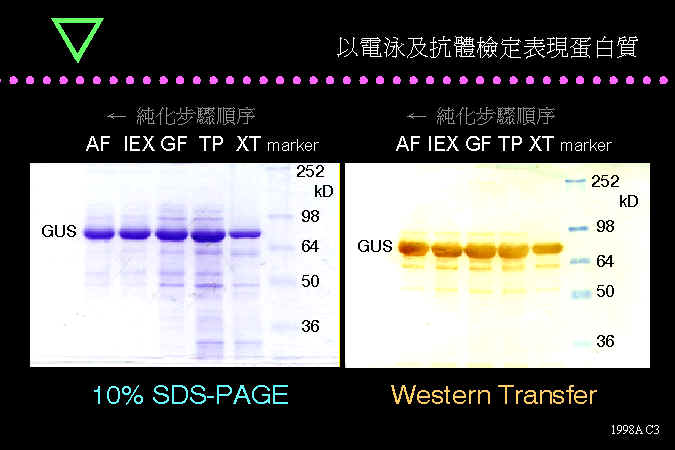
■ 以電泳及抗體檢定表現蛋白質 GUS |
||
|
圖 例 |
||
|
|
||
|
BCT 蛋白質實驗的電泳及轉印結果。 |
||
|
左圖的 Coomassie Blue 染色結果顯示,蛋白質在純化過程中,純度越來越高,最後大約只剩下 70 kD 的 GUS 色帶。平行的另一張膠片進行免疫染色,則不管在哪一個純化步驟,都染出相同的色帶,是因為抗體的專一性之故。 |
||
| A4A4 | ||
|
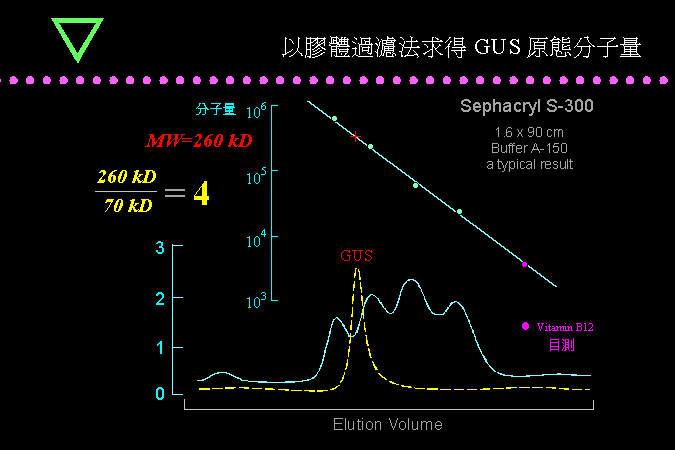
■ 以膠體過濾法求得 GUS 原態分子量 |
||
|
圖 例 |
||
|
|
||
|
比較 SDS-PAGE 及膠體過濾結果得知 GUS 是一個四元體構造。 |
||
|
膠體過濾可求得 GUS 的原態分子量約在 260 kD,而 SDS-PAGE 測得 GUS 的單元體分子量為 70 kD,因此可推得 GUS 的四級構造為四元體。 |
||
| A4A5 | ||
▼ 下一小節︰ Proteomics
|
構 造 . 純 化 . 分 析 |
|
|
|
Protein/Analysis/A4 |
|
本網頁最近修訂日期: 2001/06/30 |