圖 1 蛋白質分子間的專一性結合力量是如何構成的?
兩大類鍵結力量構成蛋白質分子間的專一性結合,其一是兩個分子之間,其構形有如積木般的互補,會因為 凡得瓦爾力 的關係,產生專一性吸引力; 另一則為兩分子之間,因為某些基團間產生了二級鍵,所造成的吸引力。通常兩個分子間的專一性吸引力的形成,這兩大類力量都會有貢獻。 親和性結合力量的大小,可以用解離常數來表示,一般解離常數至少在 10-6 以上才算有足夠的親和力 (也就是說一百萬對結合分子中,只有一對會解離開來)。
|
Biochemistry Basics |
|
|
蛋 白 質 |
|
|
蛋白質有各種催化及生理機能,是細胞的主要工作機器,其功能乃源自蛋白質分子所具有的 特定構形 及 催化活性;此種構形的形成,又因於組成蛋白質的胺基酸排列次序。 各種長短的蛋白質有不同的胺基酸組成與排列,造就了多樣而多功的蛋白質繽紛世界。 |
|
|
|
|
|
|
|
|
蛋白質構造: |
|
|
像其它巨分子一樣,蛋白質鏈也是由小分子單位 (胺基酸) 一個一個連接成的。 |
|
|
■ 細胞內巨分子的特性 (巨分子有那三種特性) |
|
|
|
|
|
一級構造: |
|
|
|
|
|
a. |
蛋白質的骨架: |
|
蛋白質的長條胺基酸序列,是為其 一級構造 (primary structure)。 此一級構造的一端為 N-端 (-NH2),另一端為 C-端 (-COOH),而以 H2N-C-C-[N-C-C]x-N-C-COOH 為骨架,其中黑體字 C 代表各胺基酸單位的 a 碳。 |
|
|
|
◆ 請練習畫出一長條胜肽的骨架,並任意加上各種胺基酸的基團。 |
|
■ 一級構造︰蛋白質的骨架 (N-C-C-N-C-C-N-C-C-N-) |
|
|
b. |
一級構造信息是由 DNA 所決定: |
|
一級構造是蛋白質最終 構形 的根本,各級構造的訊息都決定於胺基酸的序列;而胺基酸序列是根據 DNA 的核苷酸序列轉譯而來,故最終的信息是存留於細胞核的核酸中。 |
|
|
■ 一級構造︰如何定出一級序列 (有兩種方式) |
|
|
c. |
蛋白質序列可由核酸序列得知: |
|
探討一級構造主要在分析該蛋白質之 胺基酸序列,近來胺基酸序列多由該蛋白質的 cDNA 經分子群殖 (cloning) 篩選出,再經核酸定序後轉譯成胺基酸而來。 |
|
|
d. |
有三個官能基的胺基酸: |
|
除了構造之外,一段固定的胺基酸序列可能有某種特定的生理功能,稱之為 signal peptide (信息序列),同一 signal 可以在許多不同的蛋白質分子上重複出現。 例如蛋白質在 C-端若有 Lys-Asp-Glu-Leu (KDEL) 的序列,則會被回收到內質網去。 |
|
|
■ 一級構造︰序列是有意義的 (如何解讀胺基酸序列的意義) |
|
|
◆ Scientific American: A Cellular Zip Code (1999 諾貝爾生理獎 Gunther Blobel) |
|
|
|
|
|
二級構造: |
|
|
|
|
二級構造相當規律: |
|
|
|
|
|
|
◆ 請練習畫出一個胜鍵,標出胜鍵平面以及上面的各個原子。 |
|
■ 二級構造︰形成原因 (一推一拉組成二級構造) |
|
|
■ 二級構造︰氫鍵 (注意立體排列有 方向性 極性) |
|
|
a. |
a 螺旋: |
|
每
3.6 個胺基酸捲繞一圈,成為右手旋的螺旋構造,遇 Pro
則中止;由相鄰兩胜鍵平面所夾的角度,可以預測 |
|
|
■ 二級構造︰a helix 整理 (是如何形成的?) |
|
|
■ a helix 的性質描述 (氫鍵夾在 13 個原子之間) |
|
|
b. |
b 長帶: |
|
像彩帶般的構形,多由數條彩帶組合而成,相鄰彩帶之間以 氫鍵 接合,編成一片堅固的盾形平面。 依相鄰彩帶的 N→C 方向關係,可分為 同向 (parallel) 及 逆向 (antiparallel) 兩大類。b 長帶多由 R 基團較小的胺基酸 (如 Ala, Gly, Ser) 組成。 |
|
|
|
◆ 請練習畫出上述的各種二級構造,注意其立體構形及氫鍵位置。 |
|
■ 二級構造︰b sheet, turns (也是由氫鍵構成) |
|
|
◆ Protein Architecture (CMU 分子模型站,很詳細剖析 G-protein 構造,需要 Chime) |
|
|
|
|
|
◆ Alpha Helix & Beta Sheet (四個代表性蛋白質的構造分析,需要 Chime) |
|
|
◆ The Beta Helix of P. aeruginosa Alkaline Protease (最近發現由 b sheet 可組成 helix,需要 Chime) |
|
|
c. |
其它螺旋構造: |
|
除了 a 螺旋外,蛋白質也可以其它方式摺疊成螺旋構造,但每一單位螺旋的胺基酸數目不同。 當連續有數個 Pro (如 PVPAPIPP) 時,會捲成稱為 polyproline 的螺旋,每三個胺基酸轉一圈,橫切面是一個正三角形。 |
|
連結性二級構造: |
|
|
a. |
Turn 轉折: |
|
連接 a 螺旋或 b 長帶時,胜肽鏈需做劇烈的轉折,以接近 180 度的方式摺返,這些轉折點稱為 turns;其中 b turn 由四個胺基酸組成 (多含有 Gly); g turn 則由三個胺基組成,且由一個 Pro 造成主要轉折。 Turns 多分佈在蛋白質的表面,也很容易誘生抗體。 |
|
|
b. |
不規則形: |
|
除上述構造外,尚有構形不規則的連結片段,稱為 不規則形;而在蛋白質分子兩個端點附近的胜肽,活動性較大,形狀經常變化,則為 任意形 (random coil)。 |
|
|
■ 二級構造︰b sheet, turns (也是由氫鍵構成) |
|
三級構造: |
|
|
|
|
三級構造的組成力量: |
|
|
a. |
二級鍵組合三級構造: |
|
分子內各部分的二級構造再相互組合,構成完整球形的 三級構造 (tertiary structure);其構成的作用力有 離子鍵、氫鍵、疏水鍵、金屬離子 等作用力;其中 疏水鍵 對水溶性蛋白質三級構造之穩定性,貢獻最大。 |
|
|
■ 三級構造的組成力量 (四種鍵結力量 有共價鍵 -S-S-) |
|
|
◆ Bonding and Protein Structure (OMM Exhibits 顯示蛋白質 chymotrypsin 內部的鍵結) |
|
|
◆ Noncovalent Interactions in Proteins (蛋白質內的非共價鍵結) Myohemerythrin |
|
|
b. |
疏水鍵穩定蛋白質核心: |
|
水溶性蛋白質的核心緊密,多由疏水性胺基酸組成。 由於疏水性胺基酸為外界水環境所排斥,可以穩定地包埋在分子內部,維持蛋白質的完整三級構造。 |
|
|
c. |
雙硫鍵加強構造: |
|
蛋白質分子內兩個 Cys 上的 -SH 可經氧化而成 雙硫鍵 (-S-S-),雙硫鍵可加強蛋白質的立體構造。 在細胞中,雙硫鍵的形成可能需要靠酵素的催化。 |
|
◆ Amino Acid Quizzer (U Mass 的教學軟體,在一個蛋白質中指定某胺基酸,相當有趣) |
|
|
三級構造的立體構成: |
|
|
a. |
Domain 的形成: |
|
某些二級構造經常會聚在一起,例如 aaaa, bab 或 a8b8 筒狀構造等,都可自形成一小區域,稱為 domain (功能區塊),若干 domains 再組成一完整蛋白質的三級構造。 小的蛋白質通常只有一個 domain,較大蛋白質則含有兩個以上的 domains。 經常重複出現的 domain,可能有特定功用,可視為一種二級構造的再現單位 (motif)。 |
|
|
■ 三級構造: Domain 功能區塊 (介於二級與三級間的構造) |
|
|
b. |
蛋白質構形的資訊儲藏在胺基酸序列中: |
|
建構蛋白質最終立體構形的藍圖,其資訊是貯藏在一級構造的胺基酸序列中;因此某些蛋白質 (如 RNase) 在 變性 後,仍然可以回復原來的立體構形 (原態 native),稱為 復性。 |
|
|
■ 三級構造︰各種例子 (RNase, myoglobin) |
|
|
c. |
Chaperone 幫助摺疊正確構形: |
|
並非所有的變性蛋白質都可如 RNase 般復性回原態,細胞內有一類巨大分子稱為 chaperone,可以幫助蛋白質正確地摺疊成原態分子。 |
|
|
c. |
以胺基酸序列只能正確預測二級構造: |
|
若已知某蛋白質的胺基酸序列,則可以電腦運算預知某段是 a 螺旋、b 長帶或是轉折等二級構造,相當準確。 但更複雜的三級立體構造,目前則較不容易預測準確。 |
|
◆ Structure Prediction Flowchart (以流程圖列出蛋白質構造的預測方法) |
|
|
◆ The PredictProtein Server (EMBL 的一個蛋白質構造預測中心) |
|
|
◆ ExPASy: Proteomics Tools, Secondary Structure (瑞士蛋白質分析專家系統,台灣 mirror site) |
|
|
三級構造的修飾: |
|
|
a. |
蛋白質再經修飾: |
|
很多蛋白質的三級構造,即為獨立而具有活性的分子; 但有些則要再加上 輔脢、輔因子 (如 金屬離子) 或 輔基 (prosthetic group, 如 肌紅蛋白 中的 heme);有些則要再修飾以糖分子成為 醣蛋白 (glycoprotein);脂蛋白 (lipoprotein) 要連接脂質;更有的要再與其它相同或不同的蛋白質分子結合,形成四級構造 (如 血紅蛋白)。 |
|
|
■ 蛋白質轉譯後有很多修飾方式 (為何要再修飾?) |
|
|
b. |
有些蛋白質以裂解產生活性: |
|
有些蛋白質的胺基酸鏈要先經過切斷或刪除某段胜肽後,才能有活性 (如 胰島素)。胰島素是由一條基因轉譯出來的,切成三段後,其中兩段以雙硫鍵結合成具活性的三級構造,並非四級構造;這是蛋白質活性的調控方式之一。 |
|
|
◆ Insulin: Structure of a Protein Hormone (Cabrillo College 的 C4 Chemistry 教學站,需要 Netscape + Chime) |
|
|
◆ Chymotrypsin: A Serine Protease (Stryer Biochemistry 試用網路版,僅供 Netscape + Chime) |
|
四級構造: |
|
|
|
|
|
◆ ExPASy: Tertiary Structure (瑞士的蛋白質分析專家系統) |
|
|
■ 血中氧分子的運送 (Hb 如何感應血中氧分子的濃度?) ■ 下載動畫檔 Hb.pps (MS PowerPoint) |
|
|
|
|
|
a. |
次體組成四級構造: |
|
若數個相同或不同的三級構造分子,再結合成一較大的複合體,才能進行完整的活性功能,則稱為 四級構造 (quaternary structure)。 構成四級構造的每一單位分子,稱為 次體 (subunit);通常各次體之間無共價結合,而以二級鍵為主要結合力量。每一條次體蛋白質,都是由一個完整基因所轉譯出來的。 |
|
|
|
|
|
b. |
四級構造之目的: |
|
若四級構造的任一次體與受質結合之後,會誘導增強其它次體與受質之結合能力,而加速反應,則稱為 正協同作用 (positive cooperativity),反之則為 負協同作用。 四級構造在 調控 蛋白質的活性上非常重要,許多酵素是重要的例子;但典型具四級構造,且有複雜調控機制的血紅蛋白並非酵素。 |
|
|
■ 構形 (Conformation 決定所有功能性質) |
|
|
■ 蛋白質四級構造的意義 (主要是為了調節活性) |
|
|
c. |
構成性四級構造: |
|
許多病毒的蛋白質外殼,具有規律而巨大的四級構造組成。 |
|
|
■ 四級構造的作用 (調節、效率、構造) |
|
|
|
|
|
|
|
蛋白質性質: |
|
|
|
蛋白質要有正確的分子 構形,才能有效執行其生理功能;構形 或許是分子演化的基本驅策力,因為即使胺基酸序列不十分相似,同功能的蛋白質也可能有相同的 構形 (各種生物的血紅蛋白即為一例)。 |
|
■ 蛋白質的性質 (要維持其正確構形) |
|
|
變性及復性: |
|
|
|
某些條件會破壞蛋白質分子的各級構造,稱之為 變性 (denatuaration),例如 加熱、pH太 高或太低、尿素、界面活性劑、劇烈震盪等。變性的蛋白質大多會失去活性,當變性條件除去後,有些蛋白質會回復原來構形,並具原有活性,稱之為 復性。 |
|
■ 原態與變性 (蛋白質的最主要性質) |
|
|
■ SDS 破壞三級構造的疏水性核心 (一頭進入蛋白質內部 一頭在外親水) |
|
|
■ Mercaptoethanol 也是抗氧化劑 (小心使用 過多反而有害酵素) |
|
|
蛋白質構形是活動的: |
|
|
|
蛋白質分子上的各部份結構並非固定不動,而是有相當的 彈性與運動。 尤其 domain 與 domain 之間,或者酵素催化區的開閤,都有相當大的活動幅度。 這種活動會隨著溫度升高而上升,對蛋白質或酵素的活性及其調控有很大的影響。 |
|
蛋白質的專一性結合: |
|
|
|
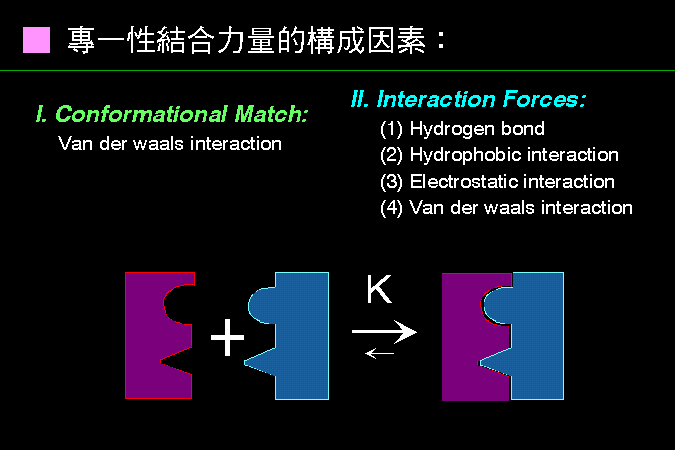
專一性在酵素的催化及細胞生理功能上,扮演重要角色。蛋白質與蛋白質之間,或與其它分子 (例如 核酸或者細胞膜),經常有專一性的結合,其構成力量如下: |
|
a. |
構形互補 (conformational match):兩分子間的結合表面,其形狀互補,像拼圖積木。 |
|
b. |
二級鍵吸引力 (interaction forces): 兩分子之結合面上,對應胺基酸間的吸引力量,由二級鍵構成。 圖 1 是一假想圖例,說明某酵素與其抑制因子間,如何進行專一性的結合。 |
|
|
|
|
|
|
|
|
|
|
圖 1 蛋白質分子間的專一性結合力量是如何構成的? |
|
兩大類鍵結力量構成蛋白質分子間的專一性結合,其一是兩個分子之間,其構形有如積木般的互補,會因為 凡得瓦爾力 的關係,產生專一性吸引力; 另一則為兩分子之間,因為某些基團間產生了二級鍵,所造成的吸引力。通常兩個分子間的專一性吸引力的形成,這兩大類力量都會有貢獻。 親和性結合力量的大小,可以用解離常數來表示,一般解離常數至少在 10-6 以上才算有足夠的親和力 (也就是說一百萬對結合分子中,只有一對會解離開來)。 |
|
|
|
|
蛋白質研究技術: |
|
|
|
蛋白質研究 通常要先純化得 均質蛋白質,然後檢定其 分子量、次體組成 及 等電點,最終則要定出蛋白質之 胺基酸序列,或其立體三次元 分子構造。 |
|
蛋白質純化技術: |
|
|
|
利用蛋白質分子量不同、表面帶電性或極性區域大小等 性質差異,可分離純化之。通常以能夠純化出大量均質蛋白質為目標,但最近的純化觀念已稍有改變,以 二次元電泳 或加上 轉印,直接挖出單一點的蛋白質,可馬上進行分析 (proteomics 的主要實驗操作)。 |
|
a. |
硫酸銨分劃法: 在蛋白質的水溶液中加入硫酸銨鹽類,會使蛋白質因疏水性區域相吸引而聚集沉澱出來,稱為 鹽析 (salting out);鹽析大略地純化出蛋白質,並可以除去核酸、醣類或脂質等物質。 |
|
b. |
膠體過濾法 (gel filtration): 依蛋白質分子量的大小不同,先後分離出來。 |
|
c. |
離子交換法 (ion exchange): 各種蛋白質的帶電性強弱不同,與離子交換介質間吸引力的大小會有差異,可以進行分離。蛋白質的帶電性,會因環境的 pH 不同而有改變。 |
|
d. |
親和層析法 (affinity chromatography): 利用分子間專一的親和性來吸引純化某蛋白質,最為直接;但並非所有蛋白質都能夠找到專一性的吸著劑。 |
|
|
|
蛋白質性質與構造檢定: |
|
|
a. |
蛋白質定量法: 染料 Coomassie Blue 與蛋白質結合後,會由褐色變為藍色; 由反應前後藍色吸光度的改變,與已知蛋白質的標準曲線比較,即可推知樣本中蛋白質的濃度;現在一般多使用 Bradford's method。 |
|
b. |
分子量測定法: 可利用 膠體過濾法、超高速離心法 (ultracentrifugation)、或 膠體電泳法 (gel electrophoresis) 來測定蛋白質的原態分子量;SDS 膠體電泳 則可測定次體分子量。 電泳 同時可以檢定蛋白質的純度如何,是解析力極佳的分析工具。 |
|
c. |
等電點 (pI): 等電焦集法 (isoelectric focusing) 極類似膠體電泳,但可測得蛋白質的等電點。 等電點是蛋白質帶電性質的重要指標,當環境的 pH 等於其等電點時,此蛋白質的淨電荷為零;大於其等電點時,淨電荷為負,反之則為正電荷。 |
|
d. |
胺基酸組成: 蛋白質以鹽酸水解成游離胺基酸,再以分析各種胺基酸之含量。 |
|
e. |
蛋白質立體構造: 以 X 光繞射法 分析蛋白質結晶,可計算出其分子的細微立體構造;近來也流行應用 核磁共振法 (NMR) 測定水溶液中蛋白質的立體構造。 |
|
f. |
質譜分析: 目前的質譜儀分析技術,已經能夠處理分子量較大的蛋白質,則可以定出蛋白質的精確分子量;甚至可以檢查該蛋白質所產生的各個片段,推出其胺基酸序列。 |
|
|
|
蛋白質序列決定法: |
|
|
|
胺基酸序列 是一個蛋白質的最根本資料,只要定出其胺基酸序列,就可以推出相當多的生化性質。 |
|
傳統胺基酸定序法: |
|
|
|
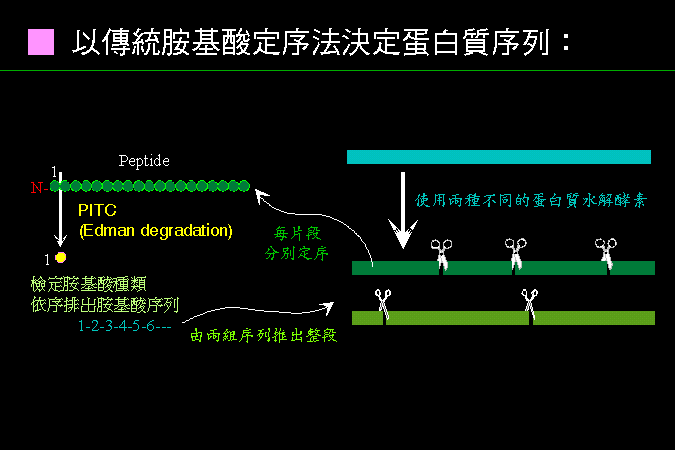
傳統的胺基酸 定序方法,是直接檢定胜肽鏈上的胺基酸種類,如圖 2 的說明: |
|
|
|
|
|
圖 2 蛋白質的胺基酸定序策略及 Edman degradation |
|
在 Edman degradation 反應中,PITC (phenylisothiocyanate) 對蛋白質的 N-端胺基酸進行修飾,產生 PTH-胺基酸,並由 N-端切開掉落出來,可用 HPLC 檢定是何種胺基酸。通常蛋白質都要以兩種水解脢分別切出兩套不同的片段,每個片段各自去定序,以便從其重疊處比對,決定各片段的順序。 |
|
|
a. |
Edman degradation: |
|
許多化學反應 (如 Edman degradation) 可由蛋白質的 N-端開始,依序一次切下一個胺基酸,再檢定每輪切下的胺基酸為何,即可推得此蛋白質的胺基酸次序。現在都用 自動定序儀器,最多可定到 50 個胺基酸。 |
|
|
b. |
蛋白質通常要先切成片段: |
|
若蛋白質太長,則無法有效定序後面的胺基酸序列;要先用 蛋白質水解脢 把目標蛋白質切成小段,各小段分別定序,然後再組合成長鏈。 |
|
|
c. |
以重疊片段排序: |
|
為了排列上述各小段胜肽的先後次序,要使用兩種不同的蛋白質水解脢,得到兩套不同長短的胜肽,分別定序後,比較各片段重疊部分,即可判斷先後次序。 |
|
|
以基因操作方法,選殖出目標酵素的 cDNA,並將其核酸序列定出,則可推出所對應的胺基酸序列;目前大都採用此種方法。 |
|
以質譜儀定序: |
|
|
|
質譜儀是利用分子的質量大小來檢定樣本,因此可以精確測出某分子的質量。若把蛋白質在質譜儀中撞擊,產生一群具有各種不同長短的片段,每一片段都剛好少一個胺基酸,然後用質譜儀一一測出這些片段的分子量,由所得各種片段分子量的差別,就可推出相差胺基酸的種類,乃至整段胺基酸的序列。 |
|
|
|
問題集 |
|
|
以下題目都沒有標準答案,許多甚至會引起很大的爭議;這樣就達到問題集之目的了。 |
|
|
1. |
血紅蛋白 (hemoglobin) 為四元体而肌紅蛋白 (myoglobin) 為單元体,請就兩者分子構造上的差異,說明其生理功能。 |
|
2. |
請問有哪些方法可以定出蛋白質或胜肽的胺基酸序列? |
|
3. |
現在 DNA 操作技術很成熟,可以把蛋白質上的某胺基酸換成別種胺基酸。 若有一個蛋白質,其分子上的某個 Leu 被換成 Val,並且某個 Lys 被換成 Arg,請問對這個蛋白質的活性影響大不大? |
|
4. |
疏水鍵 hydrophobic interaction 對蛋白質的構造有何重要貢獻? |
|
5. |
鐮形血球 sickle cell 是如何造成的? 有沒有補救或醫治的方法? |
|
6. |
構成蛋白質各級構造的各種鍵結力量中,有哪些是共價鍵? |
|
7. |
尿素 urea 可以破壞蛋白質的構造,請解釋可能的原因。 |
|
8. |
蛋白質為何需要以若干個三級構造的單元體組成一個更複雜的四級構造? |
|
9. |
氫鍵是構成蛋白質二級構造的主要力量,三級構造的組成也含有氫鍵,請問這兩類氫鍵有何不同之處? |
|
10. |
有一段蛋白質的部分胺基酸序列如下,請寫出這段蛋白質構造可能的特徵。 |
|
|
-LVRILNRILFFLWKTLTR- |
|
11. |
蛋白質構造中的 domain 是什麼? 有何重要的功用? |
|
12. |
請判斷並說明下列各題的真偽: |
|
|
a. 構成 a 螺旋及 b 長帶的主要力量都是氫鍵。 |
|
|
b. 氫鍵一定要以氫原子居中架橋。 |
|
|
c. 構成三級構造的主要力量是氫鍵、疏水鍵、雙硫鍵、離子鍵。 |
|
|
d. 構成四級構造的力量是雙硫鍵。 |
|
|
e. 蛋白質四級構造的存在,只是為了數個蛋白質分子可以聚集在一起。 |
|
|
f. Sanger 發明了 DNA 定序方法;而 Pauling 發現 double helix。 |
|
g. 每種蛋白質都有其特定構形,而構形是根基於其一級胺基酸序列。 |
|
|
h. 蛋白質的三級立體構造完全可以由一級構造推算得之。 |
|
|
i. 解讀蛋白質的胺基酸序列,可得知序列與分子功能間的對應關係。 |
|
|
j. 不規則 (irregular) 構造多在分子兩端,沒有固定形狀,有很大自由度。 |
|
|
k. 所有具有功能性的蛋白質都是由一條完整胺基酸長鏈所構成的。 |
|
|
l. 蛋白質長鏈上的 a 螺旋構造遇到 Pro 就會中止。 |
|
|
m. 各種生物血紅蛋白 (hemoglobin) 的胺基酸序列同質性很高,因此其構形相似,保有相同的運氧功能。 |
|
|
n. 水溶性蛋白質的疏水性胺基酸大都在分子的裡面,故分子內部不可有親水性胺基酸,以防分子變性。 |
|
|
o. 二級鍵的力量太弱,很難對分子間的專一性吸引力做出實質的貢獻。 |
|
|
p. Domain 可重複出現在不同分子構造,這是在基因層次已安排好的。 |
|
|
q. 所有蛋白質在變性後,都可因變性原因的去除,而恢復原態,稱為復性。 |
|
|
r. 加熱煮沸可以使蛋白質完全變性,包括雙硫鍵的斷裂。 |
|
|
s. 尿素 (urea) 可致蛋白質變性的原因是破壞 a 螺旋的氫鍵。 |
|
|
t. 一條 pI 為 5.2 的蛋白質,在細胞內可帶淨負電荷。 |
|
|
13. |
是非選擇題 (答案寫在 □ 內,是→○、非→×) |
|
1) 有關 a helix的性質: |
|
|
□ 我們可以由胺基酸的序列預測 a helix 的形成 □ 是由 Watson 及 Crick 所發現 □ 最常見的 a13 helix 是每 13 個胺基酸轉一圈 □ a helix 遇到 Pro 則中止 |
|
|
2) 蛋白質二級構造的成因: |
|
|
□ 胜肽鍵是平面構造 □ 胜鍵前後 R 基團的大小限制 □ 胜肽鏈的氫鍵吸引力 □ 胜鍵前後 R 基團的電荷限制 |
|
|
3) 有關蛋白質四級構造的性質: |
|
|
□ 有些蛋白質沒有四級構造 □ 四級構造是以共價鍵連結的 □ 有四級構造的蛋白質都具有調節作用 □ 血紅蛋白是同質四元体 |
|
|
4) 蛋白質的胺基酸序列: |
|
|
□ 可決定最後蛋白質的構形 □ 可由 DNA 的任何一股譯出 □ 並不能顯示兩相關蛋白質間的演化關係之遠近 |
|
|
14. |
構成蛋白質三級構造的力量: 1____________ 2____________ 3___________ |
|
15. |
請寫出六個以上使蛋白質變性的方法。 |
|
16. |
請說明以下人名: Sanger Anfinsen Pauling Edman Kendrew & Perutz 李卓皓 |
|
17. |
鑲在細胞膜上的許多蛋白質,其構造都有相當的相似性,多數由若干條 a helix 來回穿梭細胞膜而組成的。 請問為何多採用 a helix? |
|
18. |
在生物體內,兩個細胞之間經常要進行訊息的傳遞,例如兩個神經細胞之間,是以神經傳導物質做為神經衝動的傳遞媒介。 除此之外,兩個細胞可以利用蛋白質做為細胞間連絡的傳遞物質;其信息的傳遞,是由一個細胞傳到另一個特定的目標細胞。 而此等細胞之間,可能距離相當遠 (例如由腦部傳到胰臟),也可能有直接的接觸 (如免疫細胞間的辨認);請就蛋白質構造上的特性,說明這兩種具有專一性的細胞信息傳遞及接收方式,是如何達成的? |
|
|
|
|
|
[1] 蛋白質構造 一級 二級 三級 四級 [2] 蛋白質性質 [3] 蛋白質研究技術 [Q] 問題集
建立日期:2001/1/20 更新日期:2005/09/18 © 版權所有