|
a.
|
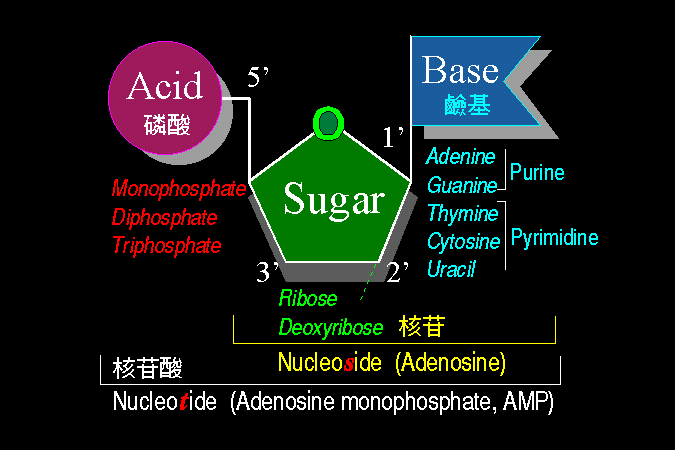
核苷酸
(nucleotide) 由三部分構成: (磷酸)- 5'
[五碳糖] 1'-{鹼基} (見 圖 1) |
|
NF1 |
|
| |
 |
| |
圖
1
核苷酸的分子組成 |
| |
核苷酸
是由三部份構造所組成,每一部份又可有數種選擇;例如五碳糖部分,可能是核糖 (ribose) 或是去氧核糖 (deoxyribose),此種差別造成了 DNA 與
RNA 在性質與構造上的不同。 我們所常見的 ATP
分子,是含有核糖,若核糖改成去氧核糖,則寫成 dATP。上圖只是畫出大略的構造關係,請自行在課本上找出確實的分子構造,要能自己默寫出詳細構造才可以;尤其各種鹼基之間,以氫鍵連結的配對情形,一定要十分清楚。
生命的奧祕,幾乎盡藏於此。 |
| |
|
| |
核苷酸除去磷酸後成為
核苷 (nucleoside):
[五碳糖] 1'-{鹼基} |
| |
(1)
五碳糖 可以是 核糖
(ribose) 或者是 去氧核糖 (deoxyribose),造成
DNA 與 RNA 的差別。 |
| |
(2)
鹼基 分成 purine (A, G) 及 pyrimidine
(T, C, U) 兩大類,T 與 U 極相似。 |
|
|
(3)
核苷 的核糖 (五號碳上)
可接一至三個磷酸,成為核苷酸,如 AMP, ADP 或 dATP。 |
| |
■
象形文字 [核 (鹼基) -
苷 (五碳糖) -
酸 (磷酸)] |
| |
◆
B-Form
DNA (完全解析一條 DNA 的立體構造,要 Chime) |
| |
◆
DNA Structure
(UMass 的 DNA
構造教學整體目錄,要 Chime) |
|
b.
|
核酸: |
|
|
前一個核苷酸的
3'-OH 端,與次一核苷酸的 5'-磷酸反應,以磷酸二酯鍵結合,接成巨分子核酸。
其中五碳糖為核糖者,即為核糖核酸 RNA;若為去氧核糖,則為去氧核糖核酸
DNA。一般而言 DNA 為雙股核酸長鏈;RNA 多為單股。 DNA
分子中,A 的數目必等於 T;而 G 數等於 C,稱為 Chargaff
定律。 |
| |
■ Nucleotide 單位以雙磷酯鍵連結
[Nucleotide 3'-(OH) → 5'-(PO4)] |
| |
◆
Base
pairs, H-bonds (UMass 的 DNA
構造教學,本圖特別強調鹼基配對,要 Chime) |
|
|
(1)
雙螺旋的一股是 5'→3' 方向,另一股則以 3'→5'
方向與之互補。 |
| |
■
核酸長鏈的寫法:有方向性
(注意 5'→3'
方向與磷、醣、鹼位置) |
| |
■
兩股核酸的方向相反 (像馬路的雙線有一定遵循方向) |
| |
◆
Ends,
Anti-parrel (UMass 的 DNA
構造教學,本圖特指出方向性,要 Chime) |
|
|
(2)
雙螺旋分子呈不對稱扭曲,因而產生有大的凹谷 (large
groove, 下圖 L) 及較小的凹谷 (small groove, 下圖
s);一個如此的扭曲單位,含有 10.5 鹼基對,其長度有
36 埃 (1 埃 = 0.1 nm)。 |
|
|
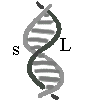 |
|
|
|
| |
■ 10.5 bp 組成一個重複單位
(有扭曲現象: grooves) |
|
|
(3)
磷酸脊骨在中性 pH 下,會帶有許多負電荷,導致兩股
DNA 相互排斥分離而變性,要加入鎂離子穩定之,是故
DNA
不能溶在純水中。真核細胞核中有帶強正電性的組織蛋白
(histone),與 DNA
結合成複雜結構,並中和掉核酸的負電性 (nucleosome)。 |
|
|
(4)
DNA 分子因其含水的多寡,可分成 A 及 B 兩型,另有 Z
型 DNA 是實驗室的產物。 |
| |
◆
DNA Structure
(OMM Exhibits
顯示 A, B, Z 三種 DNA 構造,要 Chime) |
|
c.
|
雙螺旋
(double helix): |
|
|
DNA
分子由兩股核酸捲繞而成,磷酸脊骨露在外側,鹼基在內以
A=T 及 C≡G
配對,經由氫鍵結合,兩股並相互扭曲形成雙螺旋;自然界中多屬右手旋者。 |
| |
◆ DNA
50 (BBC News 紀念發現 DNA 五十週年) |
| |
◆
The
Secondary Structure of DNA (DNA
立體構造教學,相當清楚,要 Chime) |
| |
◆ Nucleotides:
The Building Blocks of DNA
(鹼基配對的立體教學,相當清楚,要 Chime) |
| |
◆ The Double Helix
(Watson
的爭議性名著); Nature: 50 years
of DNA: Nature (1953) 171: 737 的原文 |
| |
◆ The
DNA Molecule is Shaped Like a Twisted Ladder (CSHL:雙螺旋的發現基礎
中文版) |
| |
◆ Salk
Institute
(Crick
掌管的有名研究機構); Cold Spring Harbor Laboratory
(Watson 主持的
Watson School) |
| |
◆
發現
DNA 構造的故事; Secret of
photo 51 (Public Broadcasting Service) |
|
d.
|
三級構造: |
|
|
|
|
|
 |
|
|
DNA
的長條核苷酸序列為一級構造,雙螺旋為二級構造,雙股
DNA 分子可能會捲繞成超捲曲 (supercoiling)
三級構造;這些分級都是為了說明方便,但超捲曲構造的確可以幫助大量的
DNA 擠進小小的細胞核中,而且使得 DNA
的立體構造有所變化。環狀的質體 DNA
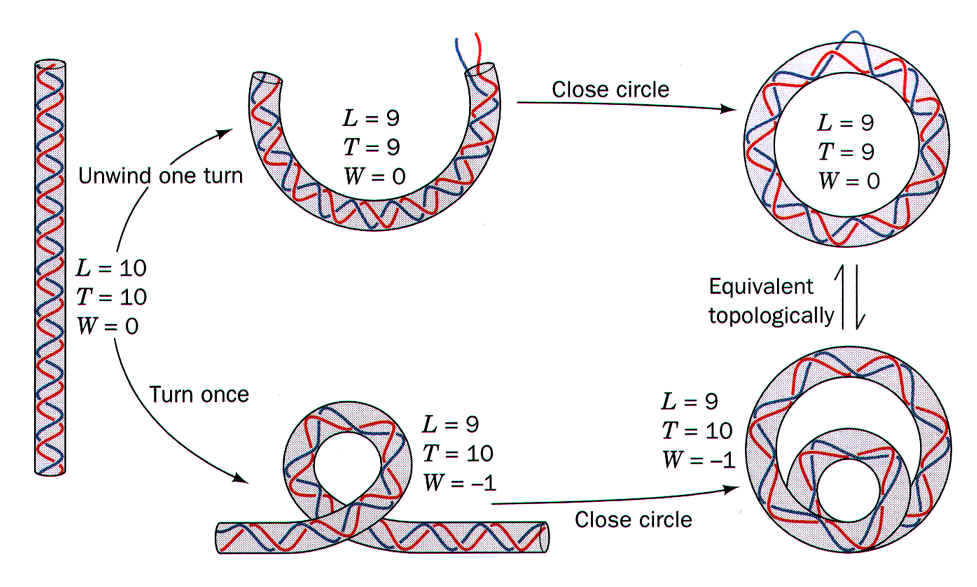
有明顯的三級構造,以下是以 105 bp
的環狀質體說明之。 請參閱下圖 Voet et al. 所著 Fundamental of
Biochemistry (1999) p.733, Figure 23-9。 |
| |
|
| |
(1)
Fig. 23-9 左圖: |
| |
雙螺旋
DNA 的兩股分子之間,以 10.5 對鹼基的長度為重覆單位,互相交叉一次,這樣的重覆單位稱為
twist (T);因此 105 bp 長的 DNA,就有 10 個交叉處 (T)。而這兩股扭曲
DNA 的立體交叉處稱為 linking,其數目稱為 linking number (L);普通的長條狀 DNA 每相互 twist 一次,自然就有一次
linking (上圖),故 T = L,10 個 T 有 10 個 L;故 L = 10, T = 10,而此
DNA 是平攤的,沒有再次捲繞,以 W (writhing) = 0 表示。 |
| |
(2)
Fig. 23-9 上排中及右圖: |
| |
但若把此段
DNA
頭尾相接捲繞成環狀,則因其分子內鍵角的緊張無法舒解,除非先解開一次扭曲
(T = 9),否則不容易形成完整的環狀構造;但因為解開了一次扭曲,因此扭曲與交叉都少一次,L
= 9, T = 9;而此環狀 DNA 還是平攤開的,故 W = 0。 |
| |
(3)
Fig. 23-9 下排中及右圖: |
| |
上述環狀
DNA 的 T 減為 9,比原來的 T = 10 少掉 1 (-1);若此環狀
DNA 要保持原來的扭曲數目 T (10),則此相差數目,必須用三級構造的超捲曲
(W) 來補足,以 L = T + W 式表之 (W = -1);也就是說,若此環狀
DNA 沒有先解開一個扭曲,則必須整個環狀 DNA
逆向捲繞一次,以便彌補之。 若 W 為正 (即 L > T),則形成
positive supercoil,為左手旋超捲曲構造;若 W
為負值,則為右手旋 (negative supercoil)。 |
| |
■ L = T + W (supercoiling 的數目)
(超捲繞可以解除核酸分子的緊張) |
| |
◆ DNA
is Packed in a Chromosome (CSHL:DNA
與 histone 的連結
中文版) |
|
e.
|
Palindrome: |
|
|
是一段特殊的鹼基序列,其特徵是在同一股
DNA 上,其 鹼基序頭尾互補,例如 GAATTC 為
EcoRI 限制脢
(restriction enzyme) 的辨認位置。
這種互補排列可能會在同一條分子內發生鹼基配對,而形成
十字型
(cruciform) 的 DNA 三級構造,可做為蛋白質辨認 DNA,或與
DNA 結合的信號;另外有利於打開 DNA (breathing),更方便上述蛋白質的辨認與結合。但蛋白質也可以不打開雙股
DNA,而直接辨認各種鹼基對的外側原子排列。 |
| |
■ Palindrome, EcoRI, sticky ends
(專一性的核酸水解酵素) |
|
f.
|
質體
(plasmid): |
|
|
DNA
多存在細胞染色體上,但許多細菌的染色體外,也有一些獨立的小分子
DNA,稱為質體。 質體是雙股環狀 DNA,常態下是以超捲曲的三級構造存在,帶有某些遺傳信息,可進出細菌菌體,是
基因操作 的重要
載體 (vector)。 |
| |
■
質體可以在宿主細菌間進出
(也帶有遺傳訊息 如抗藥性) |
| |
■
質體可接受外來的基因 (也可表現此外來基因) |
|
g.
|
RNA: |
|
|
RNA
分為信息 RNA (mRNA)、傳送 RNA (tRNA) 及核糖體 RNA (rRNA),其活動全部與蛋白質合成
(轉譯 translation) 有關。 由於 RNA 為單股分子,長條狀的脊骨活動自由,且有複雜的分子內鍵結,故分子構形較為特殊而多樣,可能具有催化活性
(ribozyme)。現今多認為地球上最早進行複製的巨分子可能是
RNA;但因為 RNA 分子構造不十分安定,後來便演化出 DNA
作為信息貯藏分子,因而造就了今日 Central Dogma
的主流。 |
| |
◆ RNA
is an Intermediary between DNA and Protein (CSHL:
如何發現 RNA 轉譯蛋白質
中文版) |
| |
◆ Some
Viruses Store Genetic Information in RNA
(CSHL:
逆向轉錄的發現 中文版) |
| |
◆ RNA
is the First Genetic Molecule (CSHL:
Stanley Miller 如何預測第一個巨分子
中文版) |
|
h.
|
基因表現: |
| |
一段基因的兩股
DNA 之中,只有其中之一可轉錄成 mRNA,這一股稱為 template
或 (-) strand,另一股則稱為 nontemplate
或 (+) strand;其表現的調控決定於此段基因之前的 DNA
序列 (promotor 或 enhancer 等)。 而 anti-sense RNA
是以人為的方法,使 (+) strand 得以轉錄出 RNA,在細胞中會與原來的
mRNA 混成,抑制該基因的表現;其中 RNA 干擾 (RNAi)
可專一性地抑制掉某個基因,且這種抑制作用可被連鎖放大。
|
| |
■ 有一面模板即可大量複製
(DNA 之一股分子上有一面模板) |
| |
■ 模板為 template (-) 股
(轉錄出來的 RNA 與 template 互補) |
| |
■ 每股 DNA 都有可能成為 template
(如何決定一段 DNA 是 template?) |
| |
■ Antisense RNA 與原 RNA 互補
(會阻止 mRNA 的表現)
|
| |
◆ DNA
Words are Three Letters Long
(CSHL:
如何解出 DNA 的三元密碼
中文版) |
| |
◆ Genes
Can be Turned On and Off (CSHL:
Operon 理論的具體說明
中文版) |
| |
◆ RNAi
- 神祕的基因糾察隊 (科學人:2003 年 9 月
p.48~56) |
| |
◆ Nature Reviews:
RNAi 動畫 (非常精彩、一定要看) |
|
a.
|
參加重要生理功能: |
|
|
核苷酸除了組成核酸外,另有下列生理功能。 |
|
|
(1)
ATP (或 GTP 等三磷核苷酸)
是攜帶能量的分子。ATP
經常會活化許多代謝小分子,以進入特定的代謝途徑;例如
Glc-1-P 被 UTP 修飾為 UDP-Glc,可參加肝糖合成。 |
|
|
(2)
構成輔脢,是某些酵素不可缺的輔助因子;如
FAD, NAD+ 及 coenzyme A (CoA)。 |
|
|
(3)
cAMP 是傳遞細胞內外信息的分子,稱為 第二傳信者
(second messenger)。 |
| |
■ 也參加其它的重要生理功能
[(1) ATP (2) 輔脢 (3) 第二傳信者] |
|
b.
|
Central
Dogma: |
|
|
Central
Dogma 敘述 DNA → RNA → 蛋白質
的流程,幾乎是所有生物體內生命現象運作的基本機制;同時
DNA 以複製來保持其自身的遺傳特性。 蛋白質合成時,tRNA
攜帶胺基酸,在核糖體依 mRNA 的信息合成蛋白質。
Central Dogma 以及基因表現的調節與控制 (基因調控),都是分子生物學的探討內容,安排在下學期的課程討論。 |
| |
■
Central Dogma
(生物學最基本的運作原則) 動
畫 |
|
|
■
生物技術學程的實驗課程 BCT
(可體驗整個 Central Dogma 過程) |
|
c.
|
變性與復性: |
|
|
DNA
的雙螺旋可因加熱而分開,稱之變性,變性後的 DNA
溶液對 260 nm 波長的吸收急劇增加,稱為 hyperchromism;
肇因於分子內的鹼基外露,而加強了吸光。若溶液的溫度再慢慢下降,則
DNA 會再回復雙螺旋的原態構造 (anneal);
回復原態的步驟,先形成一核心 nucleation (兩條單股 DNA
間的單點接觸配對),再自發地進行 zippering (由前述已結成配對的核心開始,朝兩端如拉鍊般快速拉上)。 |
| |
◆ Some
DNA does not Encode Protein (CSHL:
以變性-復性來觀察 DNA 間的互補關係
中文版) |
|
d.
|
鹼基組成的影響: |
|
|
因
G≡C 之間有三個氫鍵,A=T 間只有兩個;因此 GC
含量多的 DNA,其變性溫度 (Tm)
較高,即較不易變性;其分子也較緊密,因而密度較大。
此外,DNA
回復原態的時間,與其所含鹼基的種類、組成也都有關係。
越是複雜的 DNA,復性所需時間也越長;重複性高的 DNA
則較快;以上均可以 Cot 作圖法來表示之。 |
|
e.
|
雜合反應: |
|
|
若把兩種來源的單股
DNA 分子混合,則同質性高的 DNA 可以配對在一起,稱為
混成 或 雜合
(hybridization)。 DNA 與 RNA 之間,也可進行混成反應。 |
| |
■
復性可在不同的單股核酸間進行
(DNA/DNA, RNA/RNA, DNA/RNA) |
|
f.
|
Intron
與 exon: |
|
|
真核細胞的基因中,其
DNA 中經常插有不會表現的 DNA,稱為 intron,可能與基因的調節有關;
而基因上可以表現的部分,最後將轉錄成 mRNA,則稱為 exon。
某些 RNA 可自己進行其分子內 intron 的切除 (self splicing),具有類似酵素的功能。
這種 RNA 的 processing (加工處理),可能與基因的調控有關。 |
|
a.
|
核酸之純化: |
|
|
核酸難溶於醇類,可用乙醇或異丙醇沉澱之;
洋菜電泳可依核酸分子量的大小不同,來分離各種長度的
DNA 片段。 應用超高速離心,可分開 DNA 或 RNA
等分子密度不同的分子。 DNA
分子通常都很長,實驗操作中容易拉斷,只能得到約
100 kb 長度者。 RNA 分子較小不怕拉力,但容易受到 RNase
水解,而 RNase 很難除去。 |
|
b.
|
限制脢: |
|
|
限制脢種類很多,可在
DNA 分子上的特定鹼基序列 (一般為四或六對鹼基)
切開核酸,而此種鹼基序列,一定是 palindrome。
DNA 可能以不對稱的方式切開,得到末端不平整的 sticky ends
(或 cohesive ends);也可能平整地切成兩段,而得到鈍端 (blunt
ends)。 兩個相同的 sticky ends 可以 ligase (接合脢)
連接,是遺傳工程的基本操作。 |
| |
● EcoRI site
(GAATTC)
(限制脢可切在特定的核酸序列) |
| |
● EcoRV
site (GATATC)
(另一種限制脢) |
| |
◆
Lehninger
Biochemistry in 3D 請點選 Restrction Endonucleases (EcoRV
構造,要 Chime) |
| |
◆ The
RNA Message is Sometimes Edited
(CSHL:
介紹限制脢的作用 中文版) |
|
c.
|
核酸轉印法: |
|
|
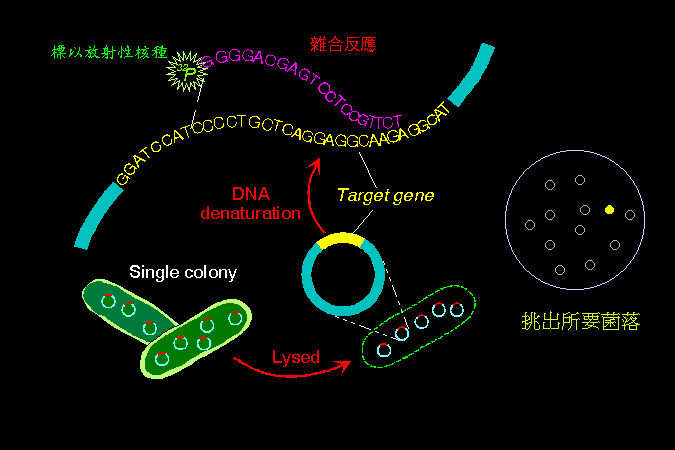
DNA
經過限制脢處理,再以電泳分離後,可轉印到 硝化纖維紙
上;然後以標有放射性的小段 DNA
為探針,進行混成雜合反應,可以挑出其中具有互補關係的
DNA 片段。 此項技術在核酸的檢定上非常重要,稱為 Southern
blotting;若用來檢定 RNA,則稱為 Northern
blotting。 探針可使用群殖或 PCR 得來的 DNA,或是化學合成之寡核苷酸片段
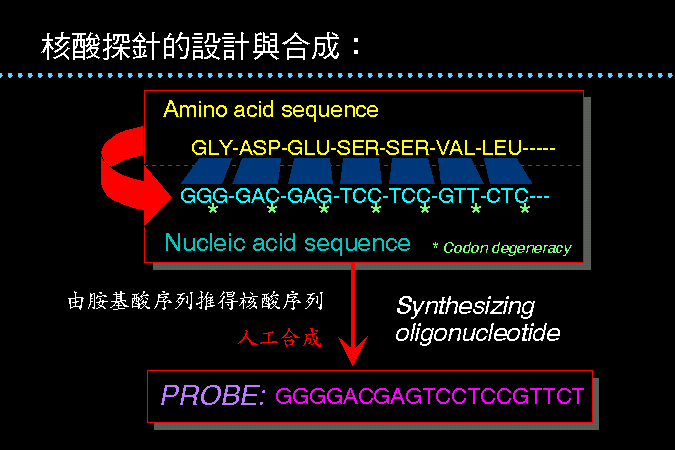
(約數十個核苷酸長度),請見 圖 2。 |
|
|
核酸雜合反應的表現極致就是
基因晶片
(gene chips) 的應用:
在約一英吋見方的小方塊中,點入數千點的核酸片段,每一點都只有一種特定核酸。
當加入樣本 DNA
與這些核酸片段進行雜合時,具有互補關係的核酸就會黏合上去,再以偵測儀器檢視樣本
DNA 可與那一種核酸片段雜合。 |
|
NF2 |
|
| |
 |
| |
 |
| |
圖
2
核酸探針的設計及應用篩選 |
|
d.
|
基因操作
(gene manipulation): |
|
|
帶有遺傳信息的
DNA 分子,可用限制脢切開後,再以 接合脢
(ligase) 接到 載體 (vector)
中;送入宿主菌後,即可大量群殖此段基因。
基因群殖 (molecular cloning) 可 放大、純化
所要的 DNA,以獲得大量且長度、組成固定的基因,以便進行此段
DNA
之定序及修飾,或研究此基因的調控特性;甚可轉殖到其他生物體,以觀察其基因表現產物。 |
| |
■
分子群殖 (molecular cloning) (可大量複製並純化出
DNA 分子) |
| |
◆『生物技術簡介』相關連結
(基因操作) |
| |
◆ Genes
Can be Moved Between Species (CSHL:
基因重組操作的最基本說明
中文版) |
| |
◆ DNA
is Only the Beginning for Understand the Human Genome (CSHL:精確修飾基因後觀察表現
中文版) |
|
e.
|
基因庫建構︰
依 DNA 來源不同,有兩種方法 |
|
|
(1)
cDNA 基因庫: |
|
|
mRNA
帶有合成蛋白質的完整信息,以 reverse transcriptase
可逆向翻製成 DNA 分子,稱為 cDNA (complementary DNA),不含
intron; cDNA 再植入載體,送入宿主中,即得 cDNA 庫。
但需注意,此種基因庫只代表正在表現中的基因,並不包括所有的基因。
cDNA 可以表現出蛋白質,並以其專一性抗體篩選之。 |
| |
■ cDNA 做法
(用逆向轉錄脢把 mRNA 轉成 cDNA) |
|
|
(2)
染色體基因庫 (genomic bank)︰ |
|
|
染色體
DNA
以限制酵素切成隨意片段後,植入載體,再送入宿主建庫。
此基因庫可能含有所有的基因,包括正在表現的,與休眠中的基因;也包含
intron,以及基因上游的調控區域 (如 promotor, enhancer 等)。
通常使用噬菌體為載體,以便容納較大的 DNA 片段。 |
| |
■ 兩種基因庫
(各有用途及特點) |
|
f.
|
PCR
(polymerase chain reaction): |
|
|
以任何
DNA (或 RNA) 為模版,加入兩段 primers 寡核苷酸,此二
primers 分別界定目標基因的起點與終點,用 DNA polymerase
往復進行複製此二 primers 之間的 DNA,則可大量合成得此段目標基因。
應用此法,可以直接群殖某特定基因,而不需先行建立基因庫,但需選擇正確的
primers。 |
| |
■ PCR
聚合脢連鎖反應
(可放大染色體中指定的一段基因) |
| |
■
一直重複下去可複製大量目標片段
(應用很廣;發明者 Mullis 已獲 諾貝爾獎) |
| |
●
PCR 複製原理 (以動畫一步一步說明
PCR 的複製機制) |
|
g.
|
DNA
定序: 兩者以 Sanger 法較常用 (見 圖
3) |
|
|
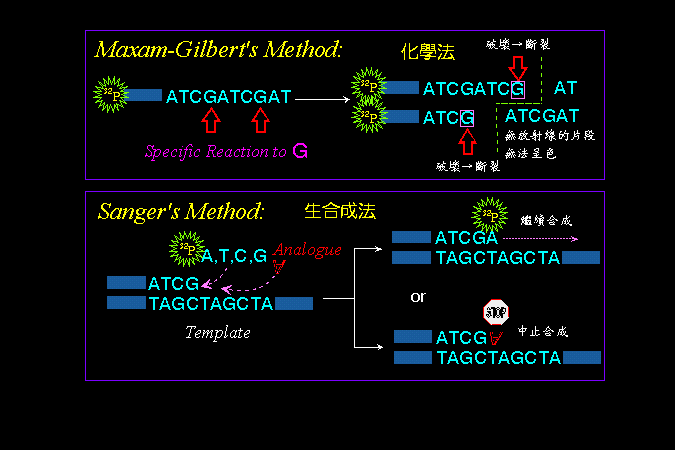
(1)
Maxam-Gilbert 法: |
|
|
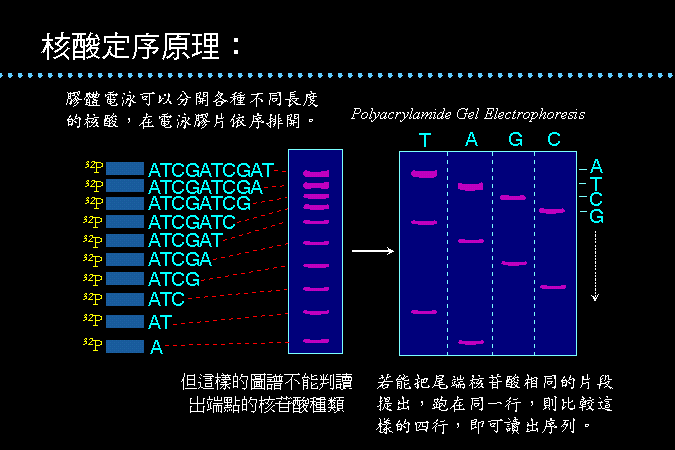
以四種化學反應分別對四種鹼基作用,每一反應只對單一種鹼基進行修飾,而在該鹼基的地方斷開,得到一系列長度不同的核酸片段。
電泳可依照這些 DNA
片段的大小,在膠體中排開,即可依序判讀 DNA
分子上核苷酸的序列;比較如此四組鹼基序列電泳,即可組合成整段
DNA。 |
|
|
(2)
Sanger 法: |
|
|
以樣本
DNA 為模板,使用 DNA polymerase 進行試管中 DNA 生合成。
四個反應中,每個反應各缺單一種核苷酸,而代以其 類似物
(analogue),則部分合成反應會停在該類似物的核苷酸處,造成各種長短不一的
DNA 片段,以電泳分離如上,即可組合判讀 DNA 序列。 |
| |
■ 雙去氧核苷酸 ddNTP
(作為 DNA 生合成反應的中止劑) |
| |
◆ A
Gene is a Discrete Sequence of DNA Nucleotids (CSHL:
介紹 Sanger 的定序生涯
中文版) |
|
|
上述兩種方法,均以
32P
標示在核酸分子上,以便顯像各不同長度的核酸片段。 |
|
NF3 |
|
| |
 |
| |
 |
| |
圖
3
核酸定序的原理及兩種定序方法的設計 |
|
h.
|
定點突變
(site-directed mutagenesis)︰ |
|
|
利用分子群殖的方法,可以改變基因上某一特定鹼基,植入載體後,在宿主中表現。
研究轉譯所得之突變蛋白質,即可獲知此特定胺基酸之改變,對蛋白質或酵素功能的影響真。 |
| |
■
定點突變 site-directed mutagenesis
(可改變蛋白質上指定的胺基酸) |
|
i.
|
RFLP
(restriction fragment length polymorphism): |
|
|
同種生物個體間的基因組成雖大致相同,但有微小差異,稱為
多形性 (polymorphism)。 這種 DNA
分子上的差異,可以用限制脢檢定出來,DNA
會被水解成不同長度的片段;進行電泳後將 DNA
轉印到紙上,再以探針偵測,比較所得圖型的異同,可得知個體間基因關係的遠近。 |
|
j.
|
人體基因序列解碼計畫
(Human Genome Project): |
|
|
2001
年已經完成人體基因 DNA
序列的解碼工作,並可由這些序列得到許多有用的資訊,此一學問稱為
genomics。
由全體基因序列可轉譯出該細胞中可能存在的全體蛋白質,稱之為
proteome,並以 proteomics (或者 function genomics)
來稱呼此一新的研究領域。 |
| |
◆ Nova:
Cracking the
code of life (有關基因解碼的完整故事) |
| |
◆ 神奇的
DNA 晶片 (科學人 2002 年 4 月 p.40~48) |
| |
◆
相關的最新技術請連結到『生物技術簡介』(基因群殖;
生物晶片) |
| |
◆
蛋白質體研究 (酵素純化與分析) |
| |
◆ A
Genome is an Entire Set of Gene (CSHL:
Watson 及 Venter 解讀人體基因密碼
中文版) |
| |
|
|
Q |
問題集 |
|
|
以下題目都沒有標準答案,許多甚至會引起很大的爭議;這樣就達到問題集之目的了。 |
|
|
|
|
1. |
以化學觀點
DNA 分子比 RNA 穩定,請說明為何?
在演化上有何作用? |
|
2. |
何為
Human Genome Project?
此大計畫對人類或科學研究,有何目的或作用? |
|
3. |
何為
palindrome sequence?
這種序列在分子的立體構造上,有何特點及作用? |
|
4. |
下列的核酸序列中,可能有若干具有意義的序列?
請畫底線標出,並加說明。 |
|
|
5'-AGGAGGATATACATGCAGAGTTAACTC-3' |
|
5. |
限制脢
BamHI 作用在 G↓GATCC 的序列,而 Bgl II
則作用在 A↓GATCT 的序列 (箭頭表示切開的位置);分別由這兩種限制脢所切得的核酸片段,混在一起以後,
能否用 ligase 接在一起?
若可以連接,則連接後可以用哪一種限制脢再度切開?
請畫出序列的剪接以便說明。 |
|
6.
|
請以對水溶解度的大小,排列以下各種核酸物質
(由大到小)︰Adenine (腺嘌呤) Adenosine (腺嘌呤核苷) Adenosine monophosphate (AMP)
Adenosine triphosphate (ATP) Deoxyadenosine。 (並請說明為何有此溶解度上
的差異) |
|
7. |
細胞內的很多反應需要蛋白質與
DNA
分子間的專一性確認與結合,例如限制脢可確認特定的鹼基序列。
但 DNA 是兩股互相纏繞的雙螺旋,鹼基對都深埋在分子內部。
請問蛋白質是如何來確認 DNA 分子上的鹼基序列?
請回答兩種以上的確認方法。 |
|
8.
|
在純化
DNA 時,最大的污染來自 RNA
及多醣類,尤其是後者很難去除之。 |
|
|
a.
如何由 DNA 中去除 RNA? 請舉三個方法。 |
|
|
b.
為何多醣類很難與 DNA 分開?
請以分子構造觀點說明之。 |
|
|
c.
如何去除雜夾在 DNA 中的多醣類? |
|
9. |
有關 DNA 的雙螺旋構造: |
|
|
a.
其分子構造中的那些因素,分別可以穩定或破壞其雙螺旋的安定性? |
|
|
b.
為何 DNA
雙螺旋構造無法像蛋白質一樣,生成具有固定構形的分子? |
|
|
c.
DNA 分子也有的三級構造 (如 super-coiling),有何生理上的功能或意義? |
|
10.
|
通常在用乙醇進行
DNA
沈澱時,要把溶液的酸鹼度調低,並且加入鎂離子。請問此種處理,有何作用?
請就 DNA 的分子構造說明之。 |
|
11.
|
基因操作技術中,genomic
bank 與 cDNA bank
在建庫的方法上及其應用上,各有不同之處,請說明之。 |
|
12. |
Central
Dogma 說明遺傳信息 DNA→RNA→Protein
的流程,三種分子均為巨分子
(macromolecule),都由單位小分子所組成。 |
|
|
a.
你認為地球上第一個出現的巨分子可能為何者? |
|
|
b.
舉出有那些實驗或結果,可證實你的觀點 (亦可自行設計實驗)。 |
|
13.
|
DNA
的雙螺旋鍊構造中,其外側為兩條帶有很強負電的磷酸脊骨,鹼基在內側以
A=T 及 C=G 的方式配對。這樣的構造,使得 DNA
分子在極低離子濃度的水溶液中,很容易變性 (denatured)。 |
|
|
a.
請說明上述導致變性的原因。 |
|
|
b.
DNA 變性後會有 hyperchromic effect 發生,請說明此現象。 |
|
|
c.
在試管中的實驗操作,要如何避免上述之變性發生? |
|
14.
|
DNA
及 RNA
為兩種遺傳上重要的大分子,其分子構造上的最大差別在於核糖分子上的一個氧原子
(2'-OH);
二級構造上,一為單股分子,另一為雙螺旋構造。因這兩點差異,導致二者在功能及性質上有截然不同的表現,請說明之。 |
|
15. |
DNA
分子上有兩股核酸序列,在進行轉錄 (transcription)
時,是使用哪一股為模版? 其決定機制如何? |
|
16. |
核酸轉譯的起始密碼只有一種,即為
AUG,可轉譯為 Met,亦即所有轉譯得蛋白質的開頭一定是
Met;但我們已發現,一般蛋白質的起頭不一定是 Met,請問為何會有這樣的結果?
並說明這種現象在細胞生理上的意義。 |
|
17.
|
進行基因操作時,若要把一段核酸送入宿主細胞時,一定要使用質體或噬菌體作為載體
(vector);請以質體為例,說明載體的功能。 |
|
18.
|
請解釋何為
intron 何為 extron? 較原始的原核細胞並無 intron,請問
intron 及 extron 是如何演化而來的? |
|
19.
|
DNA
或 RNA
等核酸構造,也會捲曲成複雜的三級構造,請舉出三例。
例如︰ 質體 DNA 的 superciol 構造。 |
|
20.
|
DNA
分子中 G 與 C 含量的多寡,會影響 DNA
的哪些分子性質? 請舉三例。 |
|
21.
|
若已知某真核細胞內的一段
RNA 序列為 -AAUAGGUACC-,則負責轉錄出此段 RNA 的 sense DNA
序列為何? 請寫出兩種可能序列。 |
|
22.
|
為何
RNA 的半衰期大都很短?
請分別就其化學性質及細胞生理學上討論之。 |
|
23.
|
請就以下各性質,分別說明能否把
DNA 及 RNA 分離開來? |
|
|
(a)
分子量 (b) 分子密度 (c) pI (d)
對醇類的溶解度 |
|
24.
|
某些
RNA 具有酵素的作用,稱為 ribozyme;請問這些 RNA
分子如何會有催化的能力? 請盡你所知回答。 |
|
25.
|
當你得到某生物染色體
genome 全部基因的核酸序列後,你可以做什麼事? |
| |
|