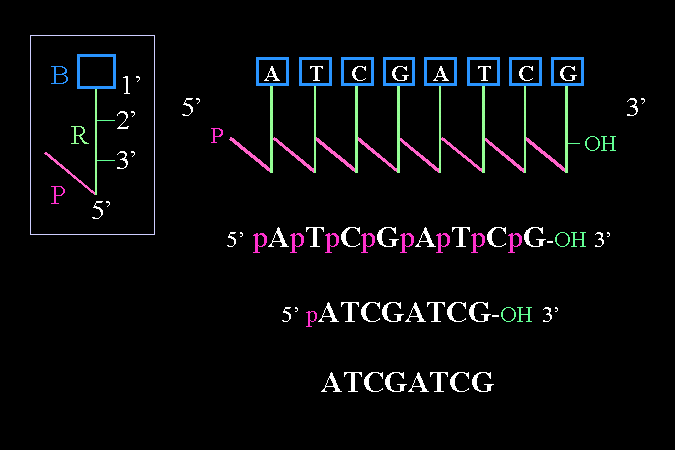核酸的構造與性質
核酸也是先把構造說明清楚後,才能對其性質有所瞭解。
■ 到講義相關部份
|
Biochemistry Basics |
|
|
核 酸 幻燈片 |
|
|
Slides |
|
|
本頁收集『核酸』的補充幻燈片,是上課時所用的自製幻燈片,其他上課圖表取自各家課本或雜誌,因為版權問題無法在此張貼,請依照幻燈片集成的清單所註明的出處,自行找尋原來圖表。 |
|
■
|
|
||
|
|
|
|
|
核酸的構造與性質 |
||
|
核酸也是先把構造說明清楚後,才能對其性質有所瞭解。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
基本的分子構造 |
||
|
核酸是細胞內的巨分子之一種,分成 DNA (去氧核糖核酸) 及 RNA (核糖核酸) 兩大類,是由核苷酸的小單位分子所組成。同樣的,核酸也是先說明其分子構造後,再以構造為基礎,來認識其種種生理性質。 |
||
|
學習核酸的構造,首要清楚辨別巨分子的核酸,是如何以單位小分子連接起來,兩條 DNA 分子如何配接起來成為雙螺旋鏈。這幾乎是所有生命現象的核心,在五十年前 Watson 及 Crick 發現雙螺旋的構造,同時也開啟了分子生物學的蓬勃研究。目前我們只能簡短說明核酸的構造與一般性質,至於圍繞著 Central Dogma 所發生的種種分子遺傳學知識,只能留待下學期講述。 雖然如此,我們還是儘量把基因操作的一些基本原理及方法,做一個簡單而概念性的說明。 |
||
|
希望同學們在這段課程內打下良好的基礎,以便在未來面對種種複雜的分子生物學問題時,能夠清楚地對付之。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
象形文字 |
||
|
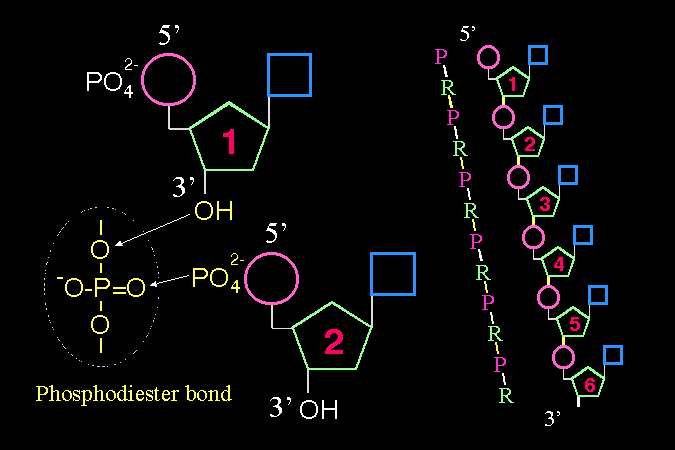
核苷酸 這三個字有點像象形文字。因為它剛好指出了 nucleotide 單位分子中的三個主要部分: 鹼基、五碳糖、磷酸。 當核苷酸去掉磷酸時,中文直接變成 核苷,英文則為 nucleoside。 |
||
|
講義上 (圖 1) 有核苷酸的分子構造,請仔細辨別各種構造上的差異,以及其所造成的影響。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
Nucleotide 單位以雙磷酯鍵連結 |
||
|
兩個核苷酸之間,前一個 (1) 以 3'-OH 與次一個 (2) 的 5'-磷酸,進行連結形成 磷酸二酯鍵 (phosphodiester bond)。 如此一直接下去,就成為核酸的長鏈。但最後有一個磷酸及一個 -OH 基是不被修飾的,因此核酸的長鏈具有方向性,可以寫成 5'-P → 3'-OH 兩端,就像蛋白質有 N-端及 C-端一樣。 |
||
|
又如同蛋白質的 N-C-C-N-C-C-N-C 骨架,核酸長鏈也有分子脊骨,它是以磷酸-核糖輪流出現的 P-R-P-R-P-R 骨架,同時在核糖分子上一個一個加上各種鹼基,組成種種序列。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
核酸長鏈的寫法:有方向性 |
||
|
核酸長鏈的記錄法,當然不能每次都把整個構造寫出來。方便的筆記法是把核糖變成一直線 (R),從上到下分別是 1' → 5',而 1' 接有鹼基 (B),5' 接磷酸 (P);各核苷酸間以斜線表示磷酸二酯鍵。 |
||
|
後來覺得這樣還是很麻煩,就省去直線及斜線,以 p 代替雙磷酯鍵,只留下各個不同的鹼基序列,最後又去掉鹼基間的 p;有時特別標出 5' 及 3' 端作為提醒,因為核酸的方向性極為重要。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
兩股核酸的方向相反 |
||
|
兩條 DNA 核酸配對在一起時,其 5'-3' 方向剛好相反;這兩股 DNA 互相捲繞成為有名的雙螺旋構造。 這種方向性,對核酸的複製及轉錄很重要,因為這些活動都有一定的方向。 |
||
|
A=T 與 C≡G 間是以氫鍵連結,其立體的分子構造,在很多書上都有。 雙螺旋分子有點像扭曲的樓梯,而 A=T 與 C≡G 就是水平的踏板,一階一階排列在兩條磷酸脊骨之間。 |
||
|
雙螺旋是由 Watson 及 Crick 在劍橋大學發現的,他們根據所收集的研究資料,即推出 DNA 的雙螺旋構造。這些資料如下: |
||
|
(1) DNA 是遺傳物質,其所含鹼基當中,A 的含量等於 T,C 等於 G。 |
||
|
(2) 由X光繞射分析結果得知,DNA 是螺旋狀,含有兩條磷酸脊骨;並且得知許多有關 DNA 分子空間排列的數據。 |
||
|
(3) A 與 T 之間可以用兩個氫鍵結合,C 與 G 之間有三個氫鍵。 |
||
|
整個發現過程充滿戲劇性,Watson 把它寫成偵探小說般的 The Double Helix,出版後雖然爭議不斷,但也可描繪出這個世紀大發現的過程。 |
||
|
Watson 與 Crick 所發現的雙螺旋,是稱為 B 型的水結合型 DNA,在細胞中最為常見;另外也有 A 型及 Z 型兩種,Z 型 DNA 最早只是實驗室中的人工產物。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
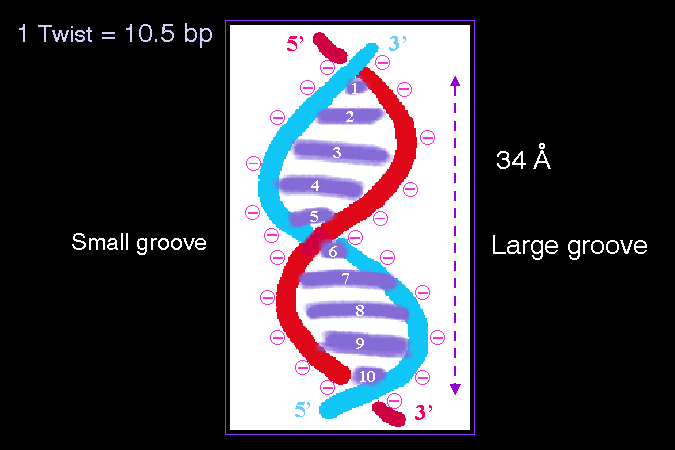
10.5 bp 組成一個重複單位有 3.6 nm 長 |
||
|
雙螺旋 是一條雙股重複扭曲的螺旋鏈,其重複出現的扭曲單位如上圖所示;以常見的 B 型 DNA 而言,每 10.5 bp 形成一個扭曲單位,長度有 3.6 nm (上圖 34A 須更正為 36A);注意每一個扭曲單位中,DNA 的一股會交過另一股一次。因為分子的扭曲並非對稱,因此分子表面會形成大小凹谷,這些凹谷對核酸與蛋白質的相互作用關係,有很大的貢獻。 |
||
|
同時因為磷酸脊骨露在外面,因此核酸的外側有相當大的負電聚集;若把 DNA 溶在純水裡,則兩股磷酸脊骨會互相排斥,造成 DNA 的兩股分開而變性沈澱,故 DNA 都要溶在含有離子的溶液中 (TE buffer)。 |
||
|
在細胞中,DNA 更會捲繞在帶有強正電的蛋白質上 (如 histone),除了抵消負電性外,還有利於 DNA 的包裝,以便容納在細胞核中。這種包裝方式,在真核細胞的染色體中,有極為複雜的組織方式。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
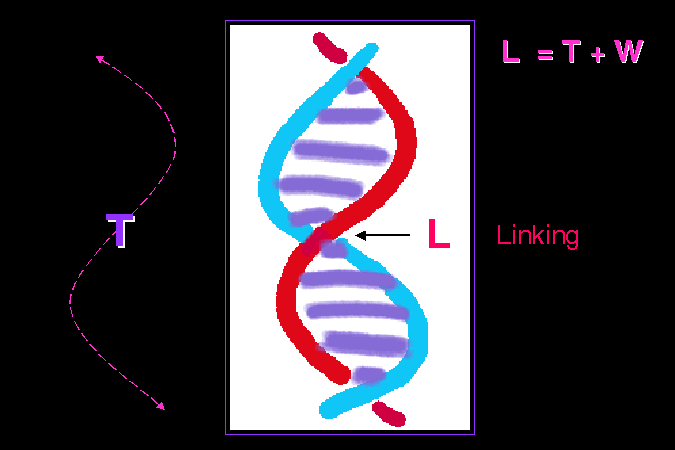
L = T + W (supercoiling 的數目) |
||
|
雙螺旋 的磷酸脊骨,雖然比起蛋白質長鏈有較大的彈性,可以較為自由地活動,但仍有一定的限制。 因此,整條雙螺旋分子並不能很自由地平攤成一直線,會以雙螺旋為軸心再度捲繞,稱為 超捲曲 (supercoil) 的構造。有點像把兩股橡皮筋以雙螺旋纏繞在一起,若把纏繞的密度加大,這條雙股橡皮筋會再度捲繞起來,成為超捲曲。超捲曲的數目 (W),與其所含鹼基數目及扭曲數目 (T) 有關,可以 L = T + W 表示之;請見講義的例子。 |
||
|
若核酸的序列為一級構造,則雙螺旋為二級構造,超捲曲則是其三級構造。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
Palindrome, Restriction Enzymes |
||
|
核酸的構成序列中,有一種特殊的短鹼基序列,其鹼基的排列是前後互補 (如 GAATTC),稱之為 palindrome;此字是文學上的用字,文句可以自由由前面向後念,或由後面向前面念者稱之 (如 madam)。 |
||
|
有一大類酵素,可以辨認核酸序列上的這種特殊構造,並且予以水解,統稱為限制脢 (restriction enzyme);這是因為含有某種限制脢的大腸菌,會限制某些入侵的病毒攻擊。 |
||
|
因為 palindrome 的核酸序列是對稱的,所組成的 DNA 構形會在長條雙螺旋上造成一個對稱的小轉折,限制脢即可認識這種對稱的轉折,並且結合到該處,再以非 Watson-Crick 的鍵結方式,在雙螺旋分子的外側凹谷 (grooves),探索其所含鹼基序列,看是否為其所能水解者。 |
||
|
較長的 palindrome 序列可以形成同一股 DNA 分子內的鹼基配對,因而形成十字形 (cruciform) 構造,兩者之間也可以互相變換;可能作為 DNA 分子上的標記位置,以便在 DNA 分子上進行各種反應。 |
||
|
|
||
|
|
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
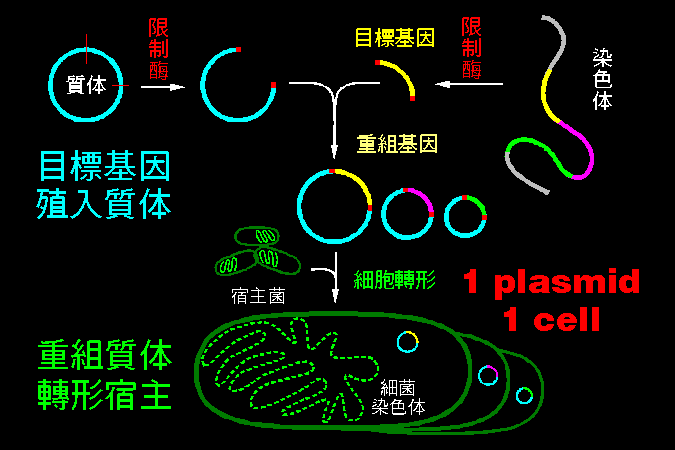
質體可以在宿主細菌間進出 |
||
|
質體是一種小型環狀 DNA,可以進出宿主細菌。 質體上面帶有可以表現出來的遺傳信息,最有名的是產生抗藥性的質體,它所轉譯出來的酵素會水解抗生素。 |
||
|
質體可以由一種宿主傳到另一種宿主菌,因此抗藥性也可以因此傳播開來,使得很多細菌很快對抗生素產生抗藥性。 遺傳工程也是利用質體的這種傳播功能,把一個被修改過的重組質體,放到宿主細菌中去表現,生產我們所要的物質。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
質體可接受外來的基因 |
||
|
質體用限制脢切開後,可以插入各種外來的 DNA,形成重組質體,後者可以轉形到宿主細胞中。 但是,一個細菌只能接受一種重組質體 (one plamid, one transformed cell),而這個轉進去宿主的質體,可以在細菌中大量複製相同的質體。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
分子群殖 (molecular cloning) |
||
|
質體在細菌中會大量複製,大約每一隻細菌可累積 5~200 個質體 (以 100 計算);若把這些轉形菌塗佈在培養皿上,則由單一細菌所形成的菌落,將只含一種轉形菌,也就是只含有一種重組質體。 |
||
|
每一個菌落若有 1000 隻細菌,而每隻細菌含有 100 個重組質體,則每個菌落就有 1×100×1000 個均質的純系重組質體,沒有雜夾其他的質體;因為一個細菌只能包容一種質體,而一個菌落只有一種細菌。 |
||
|
因此這種 分子群殖 (molecular cloning) 手法,有點像在『種植分子』,把一個目標基因,經過上述的 純化 與 放大 的過程,得到數目眾多的複製 DNA,可以抽取出來應用。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
Central Dogma |
||
|
Central Dogma 是生物界中心的基本運作法則,而其主角就是核酸,包括 DNA 及 RNA。 Central dogma 說明了遺傳信息由 DNA 流到 RNA 再到蛋白質的過程; 而其之所以能夠運作,與核酸的分子構造方式,有極大的關係。 |
||
|
DNA 的雙股構造,不但能夠說明其複製的機制,也使其信息更能妥善保存;同時也能夠複製成各種 RNA,以便行使轉譯蛋白質的功能。這一切流程,都設計得極為巧妙,而且有效。 |
||
|
RNA 雖然也是核酸,但其構造與 DNA 有很大的不同,以致於其生理功能也有相當的差異。 RNA 與 DNA 最大的不同點,在於 RNA 是單股分子,而且分子也比 DNA 小;因此,RNA 是從長條的 DNA 分子所拷貝下來的一小段功能單位,也帶有 DNA 上的密碼指令,並依此指令行事。主要的 RNA 有三大類,是為 mRNA, tRNA, rRNA 等,每個都與蛋白質的合成有關。 |
||
|
● 連結到 Central Dogma 的動畫。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
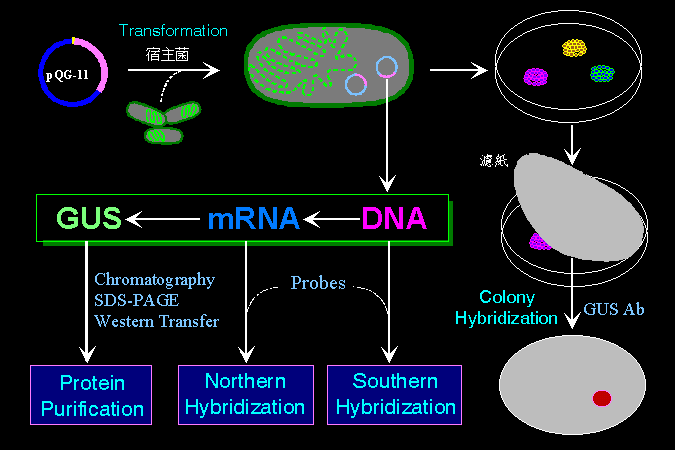
生物技術學程的實驗課程 BCT |
||
|
本校生物技術學程所開的第二階課程,都是以 Central Dogma 為對象,分別有一門講習課 (分子生物學),及一門實習課 (生物技術核心實驗 BCT)。 講習課以 Gene VII 為課本,詳細說明分子生物學上的種種活動;實習課則以 gus 基因為對象,從 DNA 純化開始,經群殖及篩選後,檢定其所生成的 RNA,並分離純化所合成的 GUS 蛋白質。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
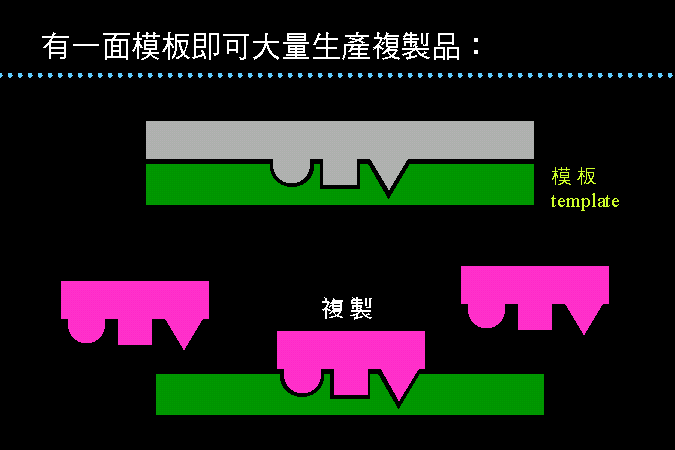
有一面模板即可大量複製 |
||
|
DNA 其中的一股好像是一個模板,打開這個模板後,就可以大量複製出互補的形狀出來。 而這個模板平時不用時,還會有一個與其互補的蓋子保護,以免模板損壞。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
模板為 template (-) 股,而 non-template (+) 那股 DNA 稱為 coding strand (2001/11/17 修正) |
||
|
兩股 DNA 中,只有一股能轉錄出 RNA,稱為 template strand 或 (-) strand;另一股則稱 nontemplate strand, (+) strand 或 coding strand。兩股上的核酸片段,都有可能成為 template,決定在於其 3' 端上游是否有轉錄起始信號。 |
||
|
RNA 的轉錄合成,一定是從 RNA 的 5' 端向 3' 端複製,而模板 DNA 則是由 DNA 的 3' 向 5' 方向讀取密碼。這些方向性一定要清楚瞭解,並且牢牢記住,因為分子生物學將由這些反應開始。 |
||
|
同時,我們一般所知的遺傳密碼 (codon),是根據所合成出來的 mRNA 來寫的;也就是說,mRNA 上面的序列就是遺傳密碼。而 mRNA 的序列是與 nontemplate (+) 相同的,而與 template (-) 互補。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
每股 DNA 都有可能成為 template |
||
|
雖然只有一股 DNA 是 template,但是兩股 DNA 的任何一股都有可能成為 template,甚至可能互相重疊。 但同一條 DNA 上的所有 templates,其閱讀方向都一樣,RNA 都由 5' 端向 3' 端複製。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
Antisense RNA 與原 RNA 互補 |
||
|
雖然只有 template strand 可以轉譯出 RNA,但也可以用人工的手法,把 nontemplate strand 選殖出來,並且送入細胞中表現。 則所轉錄出來的 antisense RNA,將會與正常的 mRNA 互補,形成 RNA/RNA 雜合分子,因而抑制了該基因的表現。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
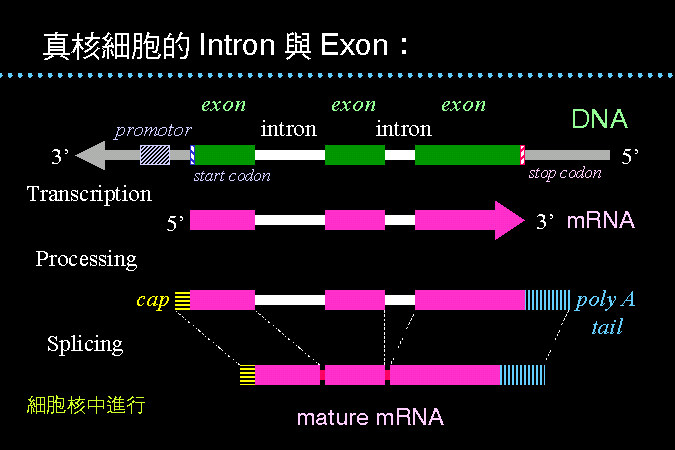
Intron 與 exon |
||
|
真核細胞 的基因構造有一點與原核細胞差別很大,就是其基因當中,插入有將來不會被表現出來的片段,稱為 intron。 當轉錄 RNA 時,這些 intron 也會如數被合成出來,形成初生 RNA,後者先經 capping 及 polyA tail 的頭尾修飾後,再以 spliceosome 切出 intron,得到成熟 mRNA,送出細胞核,以便在細胞質中轉譯蛋白質。 |
||
|
目前還不確知為何有 intron 的存在,但若以遺傳工程手法去除 intron,則此基因可能無法正常在原來的細胞核中表現。我們將以 hemoglobin 基因為例,說明 exon 與其表現蛋白質的關係。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
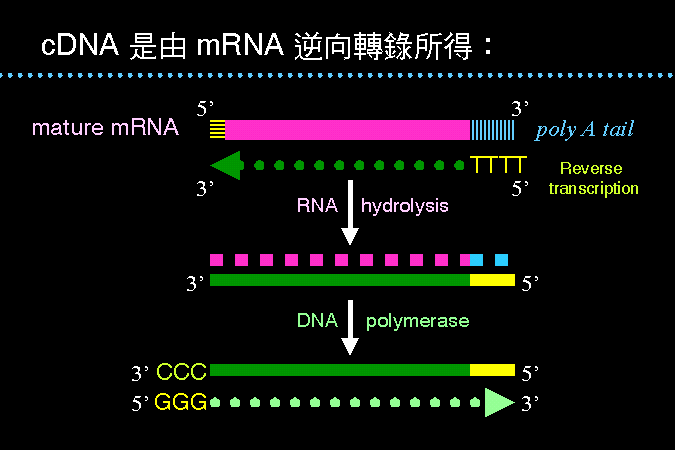
cDNA 做法 |
||
|
成熟的 mRNA 是去除 intron 的全段基因,含有完整的基因信息;因此若以 mRNA 為模板,經逆向轉錄反應所得的 DNA,稱之為互補 DNA (cDNA)。 cDNA 可以群殖入載體,並且大量表現之;以全部 mRNA 所得的 cDNA 庫,含有所有正在表現中的基因。 cDNA 選殖株可以表現出蛋白質,因此可以用抗體篩選之。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
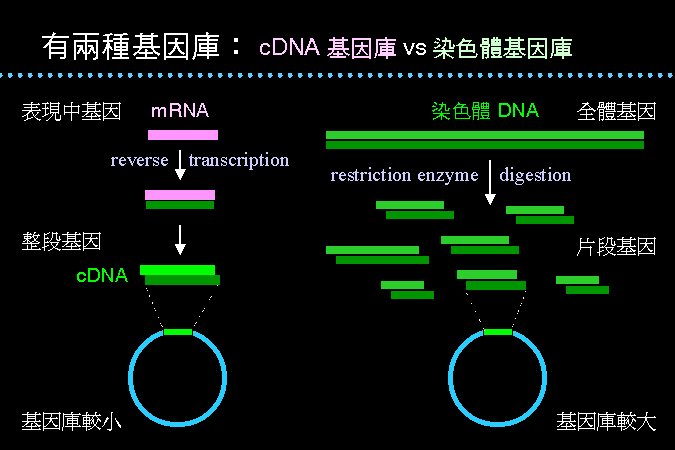
兩種基因庫 |
||
|
基因庫 的建構依基因來源的不同而有兩種方法,其一是上述由 mRNA 所得的 cDNA 來建庫 (上圖左),另一則由染色體 DNA 的限制脢片段來建庫 (上圖右)。兩者在基因庫大小與其用途上,有相當的差別,要依使用目的如何而做適當選擇。 |
||
|
許多常用的生物或細胞 (水稻),都有已經建立好的基因庫出售,只要買回來篩選,即可得到所要的基因。當然,自己必須準備好適當的探針。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
核酸的功能性質 |
||
|
DNA 的性質當然與其雙螺旋構造有關,當雙螺旋鏈解開之後,即為 DNA 分子的變性;但 DNA 分子很容易復性,回復原來的雙螺旋構造;在這一開一合之間,若加入其他的核酸分子,即可做 雜合反應 hybridization,可以檢定兩種核酸分子的序列是否相似。鹼基的組成與序列,幾乎決定了核酸分子的所有命運,因此核酸定序才成為如此重要的技術。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
也參加其它的重要生理功能 |
||
|
除了作為遺傳信息的傳遞外,核苷酸小分子也在很多的生理功能上,扮演重要角色,而且其影響範圍非常廣泛。 上面所提示的三種功能,都是所有生物都具有的基本功能,顯示核苷酸涉入演化的時間非常久遠,更加強了核酸可能在早期地球演化過程中,是最早具有演化及複製功能的巨分子。 另外,腺嘌呤 (adenine) 一直活躍地參與這些生理功能 (ATP, cAMP, NAD+),暗示腺嘌呤在早期分子演化上的主角地位。 |
||
|
|
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
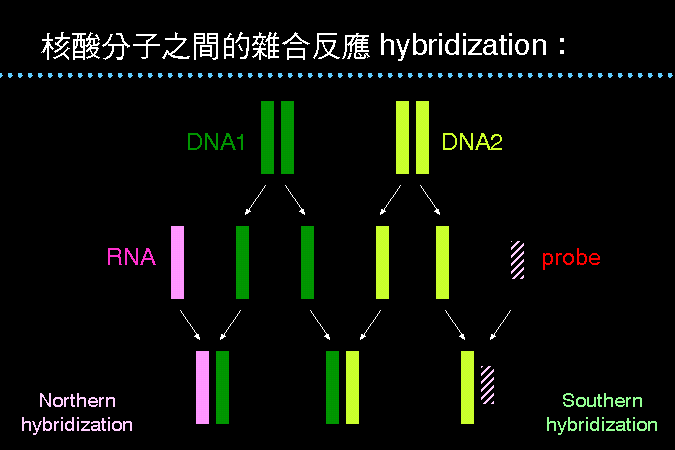
復性可在不同的單股核酸間進行 |
||
|
核酸變性之後,可以再黏合復性,回復原來的雙股核酸。若把兩種核酸經變性後混合在一起,假如兩種核酸的序列同質性高,則可能會雜合成為混成的雙股核酸。 |
||
|
也可以使用已知序列的核酸小片段,作為探針 (probe) 與 DNA 雜合;一般都先把樣本 DNA 經電泳分離後,轉印至尼龍膜上,再以放射線探針進行雜合,稱之為 Southern hybridization。 RNA 也可以在電泳後,與其互補的 DNA 序列雜合,則稱為 Northern hybridization。 |
||
|
探針的來源有很多,講義的 圖 2 舉出一個例子,說明如何由已知蛋白質的胺基酸序列回推核苷酸序列,並依此序列進行人工合成探針。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
PCR 聚合脢連鎖反應 |
||
|
PCR 技術剛被 Kary B. Mullis 發展出來的時候,居然不受重視,後來發現越來越多的重要應用;不但在研究上有貢獻,在分子診斷與法醫學上更是大放異彩,也得到 1993 諾貝爾化學獎。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
一直重複下去可複製大量目標片段 |
||
|
PCR 是聚合脢連鎖反應的縮寫,可以把指定的基因片段數量放大。 但必須知道目標基因的起始及終結的部分序列,以便分別製得界定起點與終點的兩個引子。如此,聚合脢便會在兩個引子之間複製 DNA,所得到的複製品,可在變性後再度黏合引子,進行次番複製;如此重複多次,即可得到大量介於兩個引子之間的核酸片段。 |
||
|
● 連結到 PCR 的說明動畫。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
雙去氧核苷酸 ddNTP |
||
|
核酸定序方法的精髓,在於如何製得一系列的核酸片段,這些片段各中止於不同的鹼基。 講義的 圖 3 說明這些定序方法的原理,有兩種方法可以製備一系列長度的核酸片段。 |
||
|
目前都用 Sanger 的生合成法,以目標核酸為模板,加入四種核苷酸及 polymerase,複製該段核酸;但除了所加的四種核苷酸中,額外加入某一種核苷酸的衍生物 (dideoxynucleotide),則合成反應將可能止於這種核苷酸的位置,因而得到各種長短不同的片段,以電泳可以依序排列出來。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
核酸研究技術 |
||
|
核酸的研究技術,大多已散佈在上述構造與性質的說明中,因此只補充說明未提及的部分。 |
||
|
■ 到講義相關部份 |
|
|
|
|
||
|
|
|
|
|
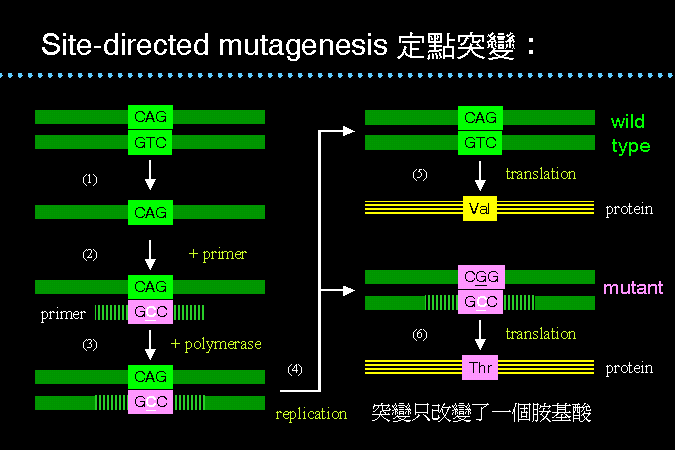
定點突變 site-directed mutagenesis |
||
|
定點突變 是使用人工合成的引子,先與目標基因雜合 (2),而此引子的核苷酸序列與目標基因互補,但其上有一個核苷酸變異 (GTC→GCC)。然後補滿雙股核酸 (3),再把此混成基因轉殖進入宿主,則宿主菌的複製系統將會因複製而產生兩種質體 (4),其一為原來的野生型,轉譯可得原來的蛋白質 (5);另一則為突變型,轉譯則得到突變的蛋白質,但只改變了一個指定位置的胺基酸 (6)。 |
||
|
■ 到講義相關部份 |
|
|
回到核酸講義 [1] 分子構造 [2] 功能性質 [3] 研究技術 [Q] 問題集 幻燈片集成 Reference
建立日期:2000/1/19 更新日期:2003/10/27 © 版權所有