
圖 10 ELISA 三明治法
|
Biochemistry Basics |
|
|
本網頁一律以『脢』表示 enzyme |
|
|
細胞代謝與酵素調控: |
|
|
|
酵素與細胞代謝極為密切,因為所有的代謝路徑,全是由酵素與其基質所組成的;此外,還要加上對酵素活性的調節控制,以便使得細胞正常運作。 |
|
■ 酵化講義目錄 第 7 節 (開車上路注意紅綠燈與號誌) |
|
|
細胞代謝途徑: |
|
|
|
|
|
代謝調控原則: |
|
|
|
|
|
a. |
一個基因一個酵素: 細胞的代謝途徑極為複雜,但並非細胞所有的代謝路徑都在進行;進行中的每一步代謝,都由某一種酵素負責催化反應,而每一個酵素分子都是由一個對應基因所轉錄。因此某基因的開啟或關閉,到最後可能會影響代謝途徑的進行。 |
|
b. |
速率決定步驟: 因此控制該酵素的 活性 或 生合成量,即可控制該步驟反應的快慢;若此反應為某一系列代謝路徑的 速率決定步驟 (rate-limiting step),則可控制這整條代謝途徑。 |
|
c. |
可逆或不可逆: 大部份酵素反應是 可逆 的,有時為了使反應保持在某一方向,則成為不可逆反應;不可逆反應大多與消耗 ATP 的反應耦合。 |
|
d. |
代謝路徑可互通: 許多代謝路徑間若有共同的中間物,則可互通,也可能有旁支或小路相連;因此若某條重要路徑失效,細胞通常不會立刻死亡,而會互補保持一種動態的穩定狀況。 |
|
◆ Metabolic Pathways (網路版代謝路徑,可以找到部分主要代謝反應) |
|
|
異化代謝途徑鳥瞰: |
|
|
以生物分子而言,異化代謝路徑可分成三個層次,有計畫地把巨分子逐步分解,最後得到能量;其中最主要的是一條 糖解作用 (glycolysis)。 |
|
|
a. |
Stage 1: 巨分子 (蛋白質, 多糖, 脂質) 消化成單元小分子 (胺基酸、單糖、脂肪酸),可說就是消化作用。 |
|
b. |
Stage 2: 單元小分子再分解成更小的 acetyl CoA,初步得到一部份的能量。 |
|
c. |
Stage 3: Acetyl CoA 經過氧化磷酸化反應得到大量能量,分解成 CO2 及水。 |
|
|
■ 7.1 先瞭解主要代謝路徑 (切牛排) (六碳糖的分解: 6 3 3 2 1 碳數減少) |
|
糖類中心代謝途徑: |
|
|
由糖解作用到氧化磷酸化反應,是細胞最主要的一條代謝大道;學習細胞的代謝途徑,可以此為中心,其它各類大小分子的代謝都可匯入此中心。 |
|
|
a. |
糖解作用: 把葡萄糖分解成 pyruvate → acetyl CoA (所有代謝分子的焦點)。 |
|
b. |
檸檬酸循環:在粒線體中把 acetyl CoA 再分解成 CO2,產生 NADH。 |
|
c. |
氧化磷酸化反應: NADH 轉換成 ATP 並生成水。 |
|
■ 主要代謝路徑整理 (Glycolysis) (如何獲得能量? 分解 → 氧化) |
|
|
代謝途徑中酵素的調控: |
|
|
|
|
|
■ 細胞如何控制酵素 (各層次的調控方式不同) |
|
|
基因表現的調控: |
|
|
|
|
|
a. |
酵素的 生合成 受到其基因的控制 (DNA→mRNA→protein),因此基因的開或關,或其表現程度,會影響該酵素在細胞內的量,進而影響該酵素所控制的代謝反應,是為 基因表現 (gene expression)。 一個表現中的基因,應該有大量 mRNA 轉錄出來,但有大量 mRNA 卻不一定產生大量酵素。 |
|
b. |
一條代謝路徑的起始基質,可能誘導關鍵酵素基因的表現,稱為 induction;此代謝的最終產物,也可能抑制酵素表現,稱為 repression。 基因操縱子 (operon) 即是一例,乳糖能夠去除 repressor 與 lac operon 之結合,而使基因開動。 |
|
|
■ 7.2 基因的表現 (有許多個控制點) |
|
酵素活性調節: |
|
|
|
|
|
a. |
|
|
|
使用 cAMP 或正負迴饋的 效應物 等方式,以非共價方式修飾酵素活性,此多為可逆性的調控。勿把異位脢與上述操縱子的調控方式混為一談,兩者都可被基質活化,但前者是修飾酵素本身,而後者是影響基因的表現。 |
|
b. |
共價修飾: |
|
以磷酸化或蛋白質水解的方式,來增強或降低酵素活性。 多為 cascade (梯瀑) 式的連鎖代謝反應,cascade 連鎖反應會放大 (amplify) 某條代謝路徑的活性。除了正向的以生合成增加酵素量之外,蛋白質的降解也是一項重要的調控方式,並以蛋白脢或 ubiquitin 配合 proteasome 進行降解,以除去此酵素。 |
|
|
激素調控: |
|
|
|
|
|
a. |
荷爾蒙 (如 insulin) 傳遞 長程 生理指令,經由血液傳送並與目標細胞接觸,結合到細胞膜上的接受器 (receptor),可引發一系列的信息傳導反應,把『啟動』的指令傳入細胞內,藉著蛋白質激脢或磷酸脢等,活化某些關鍵酵素,開始進行該細胞所預設的功能 (如分解肝糖),以應身體所需。 |
|
b. |
細胞激素 (cytokine) 則多在免疫細胞之間傳遞 短程 的信息,可刺激局部的細胞增生;也是利用信息傳導路徑,來啟動細胞內的各種生理功能。 |
|
細胞空間的效用: |
|
|
|
|
|
a. |
胞器隔離: 真核細胞內有許多 隔間 (compartmentation),形成其胞器的隔離空間;在胞器內可聚集較濃的特定酵素及基質,以便有效進行催化反應,或避免有害的副作用。 例如葡萄糖中心代謝途徑中,檸檬酸循環及氧化磷酸化反應是在粒線體中進行的。 |
|
b. |
酵素複合體: 幾個連續反應的酵素,可組合在一起成為複合體;則某酵素的生成物,可馬上被下一個酵素用為反應物,降低擴散碰撞所需時間。 例如葡萄糖中心代謝途徑中的 pyruvate dehydrogenase (催化 pyruvate → acetyl CoA) 是由三種酵素組成的複合體。 |
|
c. |
膜上酵素群: 一群連續反應的酵素,也可以一起附在細胞膜上,除了可加速反應速率外,其作用也可能與細胞膜有關。 例如 葡萄糖中心代謝途徑 中,氧化磷酸化反應酵素群 是附在粒線體的內層膜上,並利用膜內外的質子濃度差異來產生 ATP。 |
|
研究酵素及代謝的材料: |
|
|
酵素材料的來源或取得,依其不同目的可有數個層次。 |
|
|
a. |
通常我們必需純化出 均質酵素 (homogeneous enzyme),以針對某一特定酵素進行研究,因此蛋白質純化技術在生化研究上極為重要,請複習這些純化及檢定方法。 |
|
b. |
但複雜的生化反應,並非單一的純質酵素所能達成,因此可分離出某種特定細胞,或者將細胞打破所得到的 溶胞液 (cell-free lysate),以作為研究酵素或代謝反應的材料。 動物細胞大多可以建立 細胞培養株,某些植物也可以 癒創組織 (callus) 培養細胞。 若研究需要利用特定的 胞器 (organelle),則可由細胞的溶胞液經 超高速離心法 取得。 |
|
c. |
有時需要以整個生物體 (whole organism),或其部份器官 (organ) 來進行實驗,以便觀察巨觀的反應。 反應可能相當複雜,因此以放射線物質或用專一性抗體來追蹤是較清楚的方法。 |
|
d. |
不論何種層次的研究,都要注意目標酵素是在那一個 生長時期,或是那一種 器官 內表現的。 例如:抽取檸檬酸循環的某些酵素,可以先分離出粒線體來。 |
|
e. |
生物化學研究大多是以分離純化的酵素來進行研究,要小心求證所獲得的結果,是否確實為自然的生理現象,而非僅是試管中的假象。 一切觀察均得回歸整體性的細胞生理,否則難有重大意義。 |
|
◆有關酵素純化與分析方法,在生物化學實驗課程中,應該已有部分的基本操作;若要有更詳盡的技術引導,請參考研究所課程的『酵素化學實驗』。 |
|
|
■ 酵素純化方法的原理 (利用不同蛋白質性質間的差異) |
|
|
■ 如何分離這些大大小小積木? (以假想的例子說明純化方法) |
|
|
■ 酵素純化方法整理 (各種純化方法所根據的原理) |
|
|
|
|
酵素在生物技術上的應用: |
|
|
|
酵素在現代生物技術的研究或應用上,是一個非常重要的範疇。雖然基因群殖等分子生物的研究蓬勃,但基因表現的產物仍是蛋白質或酵素;另外,在基因操作所應用到的核酸剪接工具,幾乎全部是酵素。 以下列舉部份目前生物技術研究常見的一些應用,以及最近的重要發展。 |
|
|
||
|
|
||
|
|
|
|
|
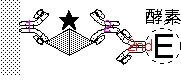
圖 10 ELISA 三明治法 |
||
|
固定化酵素及酵素電極: |
|
|
酵素經固定後有許多好處。 |
|
|
a. |
用物理或化學方法把酵素固定到固相擔體上,比一般使用的溶態酵素有以下優點: |
|
|
(1) 酵素 可回收 重複使用,較為經濟。 |
|
|
(2) 酵素的 穩定性 提高,可能較耐熱或極端的 pH。 |
|
|
(3) 固相與液相的 分離方便,使用上速度快而分離完全,有助於自動化。 |
|
|
(4) 許多酵素是附在細胞膜上,固定化酵素可 模擬 細胞內酵素的實際環境。 |
|
b. |
利用上述酵素的固定化,把酵素固定在半透性薄膜上,連接到電極,偵測反應進行的結果 (例如 pH 的改變),可作為酵素反應的自動化偵測工具。 |
|
蛋白質工程及人造酵素︰ |
|
|
以基因重組或其它方法,可以大量生產某種酵素,也能改變酵素的催化特性。 |
|
|
a. |
分子群殖 molecular cloning: |
|
|
cDNA 包含完整的蛋白質轉譯訊息,若把某蛋白的 cDNA 插在表現載体中 (如某質體),則宿主可能表現此段轉殖基因,而生合成此蛋白質。若在此 DNA 接上另一種已知的酵素基因 (如 luciferase 或 GUS),則表現出來的蛋白質是二者的連結體,稱為 融合蛋白質 (fusion protein);而此酵素的活性可做為追蹤之用,稱為 reporter。 |
|
b. |
|
|
|
若能改變酵素活性區的胺基酸,則可能改變酵素的活性,或是其專一性。 通常先研究並預測改變其活性區某胺基酸後,可能引起的變化;再以 人工定點突變 (site-directed mutagenesis) 改變某核苷酸,然後以分子群殖操作表現該突變蛋白質。 |
|
c. |
人造酵素: |
|
|
酵素的活性區通常包含數個極性胺基酸,若在人造的分子骨架上,模仿活性區的幾何位置,接上這些胺基酸,則可能得到具有催化作用的人造分子。 |
|
d. |
Abzyme (催化性抗體): |
|
若能得知酵素催化反應過程中,其基質轉換為產物的 過渡狀態 物質,以此物質或其類似物作為抗原進行免疫,則所得到的抗體,可能具有催化能力。 但其催化效率,遠不及自然酵素,通常只有千分之一的效果。 最主要原因在於酵素的催化區是一凹陷口袋,可隔離外界干擾,提供最佳環境穩定過渡狀態;而 abzyme 的結合區較淺,無法十分有效地隔離並穩定過渡狀態 (Nature 1996, 383: 23-24)。 |
|
|
Proteome 蛋白質體: |
|
|
|
|
Genome project 基因體計畫: |
|
|
|
|
|
◆ Genome Gateway (Nature 人類基因體網路專欄) |
|
|
Proteome 蛋白質體: |
|
|
a. |
一旦解出某種生物的染色體 DNA 序列,接著有許多工作可以進行,其中最有趣的是可以馬上把這些序列翻成將表現出來的蛋白質,如此我們就可以得知某生物細胞內可能含有的全體蛋白質,稱之為 proteome 蛋白質體。 |
|
|
■ Genome → Proteome (基因座的整體蛋白質表現) |
|
b. |
生物細胞內的蛋白質體中,當然只有一部份是酵素,但由細胞內所含有的酵素種類,即可拼出該細胞可能的代謝途徑;由這些代謝途徑,就可以推出某細胞是如何運作。 |
|
c. |
這種研究方式,與傳統的生物化學或生理學實在相當不同,要靠完整的資料庫與功能強大的電腦軟體,是 生物資訊學 (bioinformatics) 的主要範疇。 |
|
■ 2D 電泳 →→→ Mass 定序分析 (快速分析表現蛋白質) |
|
|
◆ Functional Genomics (Science 蛋白質體相關消息的網路專欄) |
|
◆ 生物技術簡介 (以網路連結介紹生物技術);也有文字說明:生物技術的發展與未來 |
|
|
|
建立日期:2001/3/19 更新日期:2004/06/12 © 版權所有