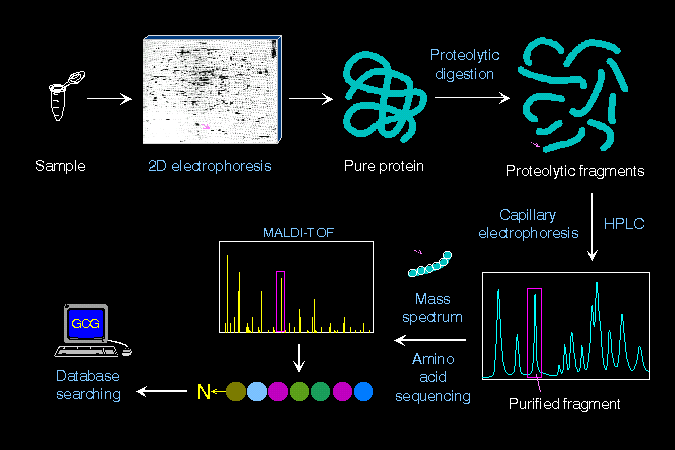
酵化講義目錄 第 7 節
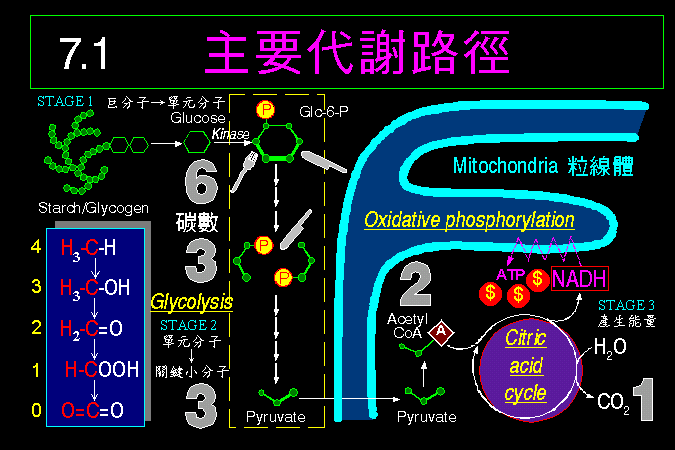
熟悉酵素的各種性質後,生化課將要進入各種代謝途徑,因為每種代謝反應,都有一個酵素在做催化。 首先我們將簡介何為代謝途徑 (7.1),說明調節代謝途徑的一些基本原則,並以異化代謝為例,說明細胞如何有計畫地把食物分子逐步分解成小分子,然後進入細胞內最重要的一條代謝大道 (糖解作用 glycolysis)。
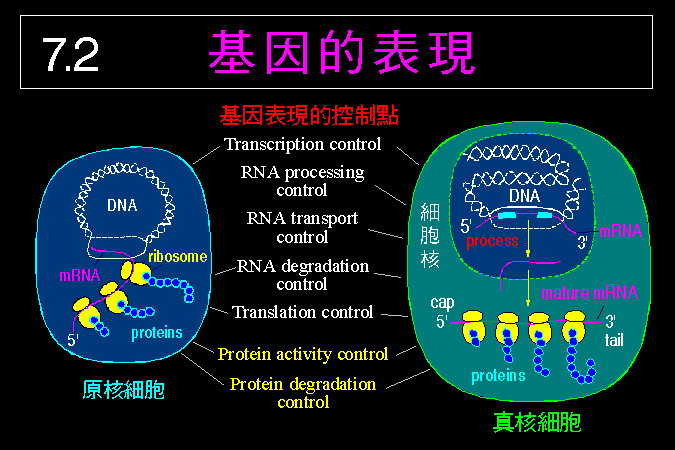
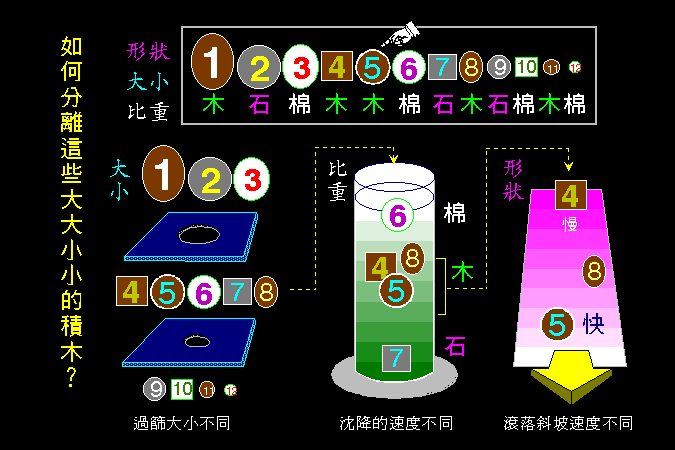
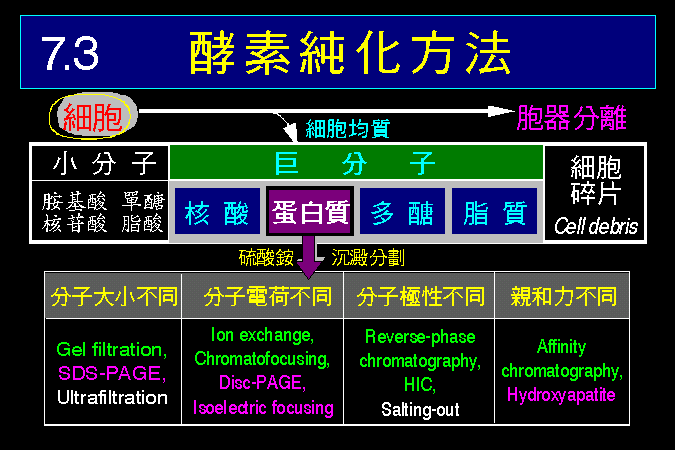
接著將整理出細胞內代謝途徑的調節方法 (7.2),如何以控制酵素的量或活性來控制代謝途徑,如何以荷爾蒙來開或關閉代謝途徑,細胞甚至可以用空間上的效應來加強酵素系統的作用。最後我們簡要介紹研究酵素及代謝系統的材料種類,以及材料的取得與處理方法 (7.3)。
這一節的討論,多相當鉅觀,沒有太多分子層次的解析。不過,這種大範圍的審視也十分重要,以免走入死胡同而不知全貌。現在的分子生物學研究,也都要回歸到整個細胞、生物體、甚或整個生態的佐證,才能更有力地證明分子層次的發現是否有意義。
■ 到講義相關部份