|
|
生物化學基礎 Biochemistry Basics 2008 酵素 Enzyme (5) |
回首頁 ■ |
|
|
|
|||
|
1 酵素的命名 2 酵素的構成 3 酵素動力學 4 酵素的抑制 5 酵素催化機制 6 酵素活性調節 7 細胞代謝調控 8 生物技術應用 |
5 酵素的催化機制 5.1 酵素活性區 5.2 協同式催化機制 5.3 順序式催化機制 5.4 酵素的專一性 5.4.1 專一性結合區 5.4.2 專一性結合力量 5.4.3 立體專一性 問題集 (5) |
Chymotrypsin 的奇妙催化機制 |
|


|
■ 酵素 5 |
■ 相關投影片 |
||||
5 酵素動力學:酵素的催化機制可以化學反應逐步推得,與分子的構造也有相當大的關係。
催化反應都在活性區內進行,活性區可提供有利催化的環境。 a. 活性區:是酵素與其基質 (或輔脢) 的結合區,並在此進行催化反應。活性區通常是一凹陷袋狀構造,水分子不易進入袋中。活性區內的胺基酸,只有那些具反應性基團 (reactive group) 者,才直接參與催化;但各種胺基酸都有可能參與結合基質。 b. 酵素催化的化學機制,通常是以下面三種基本方式進行反應: (1) Bond strain:基質結合到酵素後,酵素的構形扭曲,使基質分子內某共價鍵受力斷裂。 (2) Acid-base:利用活性區內胺基酸殘基或輔脢,可以放出或接受質子 (或電子) 的特性,對基質進行質子或電子的轉送。 (3) Orientation:活性區可提供基質適當的排列空間,使有利於反應進行。 以上三種方式或可同時發生,為協同式 (concerted set);亦可先後依序發生,稱順序式 (sequential mechanism)。以下兩個重要例子都是蛋白脢。
|
|
||||
|
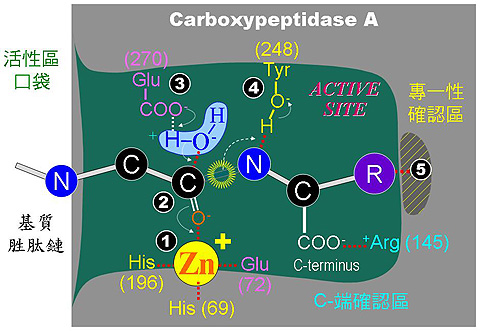
5.2 協同式催化機制: 以 carboxypeptidase A (CPA, 羧肽脢) 為模型酵素說明 (圖 3)。 a. CPA 的分子量 34 kD,含 307 個胺基酸,有一雙硫鍵,及一個鋅離子,是一種金屬蛋白脢。 CPA 可由胜肽的C-端,依序切下外側胺基酸 (外切脢),當胺基酸的 R 基團為非極性者,較有利反應進行;而 carboxypeptidase B (CPB) 只切 C-端為 Lys 或 Arg 者,二者專一性不同。
圖 3 Carboxypeptidase A 的催化機制 ◆ Carboxypeptidase 催化機制 (以動畫呈現上圖之催化動作) b. CPA 的詳細催化機制如圖 3 所示,並加說明於下: (括號內數字與圖 3 中對應) (1) Zn2+ 離子乃重要輔助因子,可吸住基質胜肽鍵上的 carbonyl 基,增強其極性,使 a 碳帶正電。 (3) Glu270 吸住水分子,放出 OH- 攻擊 C+ (2),產生新的 C-OH 鍵。 (4) Tyr248-OH 上的質子,與氮 lone pair 電子產生新鍵,原來的胜鍵斷裂。 (5) 附近的胺基酸與基質 C-端的 R 基團,有專一性的結合,以辨別基質的極性;同時 Arg145 與基質 C-端的 -COOH 結合,確定基質蛋白質是以 C-端進入活性區。
|
|
||||
|
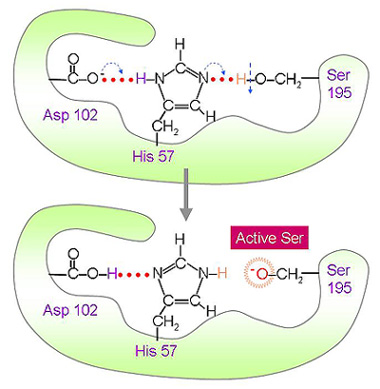
5.3 順序式催化機制: 以 chymotrypsin (CT, 胰凝乳蛋白脢) 為代表,請找出課本的相關圖片。 a. 分子構造:CT 的分子量為 25 kD,含三條胜肽,由兩個雙硫鍵連接,是轉譯後修飾的產物;剛轉譯出來的完整胜肽鏈沒有活性,要在接近 N-端的 Arg15 與 Ile16 之間先斷開後,CT 才能活化。 b. 催化活性:CT 可水解胜肽鏈上面的芳香族胺基酸 (Tyr, Phe, Trp) 或 Met (具較大非極性基團者),切開這些胺基酸 C-側的胜鍵,是一種內切脢。 c. 活性區:CT 大部分的極性胺基酸都露在分子外表,只有三個在活性區內,對催化反應扮重要角色 (Ser195, His57, Asp102)。三者以『電荷接力』形成高反應性 Ser,其 Ser195-OH 基上的 H+ 被鄰近的 His57 吸收,生成具有高反應性的 -O -。 ◆ Chymotrypsin 活性區鐵三角 (以動畫呈現活性區之生成)
圖 4 Ser 蛋白脢催化區上由三個胺基酸所構成的電荷接力 d. 催化機制:Ser上的高反應性 -O- 會攻擊基質胜肽上的 carbonyl 碳 (帶微正電),形成新的 C-O 鍵 (acylation 步驟),同時斷開基質的胜鍵,先釋出一段 C-側的胜肽;然後加水分解,破壞此 C-O 鍵 (deacylation 步驟),釋出另一段 N-側胜肽。以上兩個步驟,依次順序發生。 ◆ Chymotrypsin 催化機制 (以動畫呈現上述之催化動作) e. 穩定過渡狀態:除了 catalytic triad 可以有效催化之外,活性區也可穩定催化反應的中間過渡狀態。過渡狀態有相當高的負電荷,因此以活性區附近的 Gly193 及 Ser195 本身的 -N-H 與之產生氫鍵而中和之。 f. Ser 蛋白脢家族:這種以 Ser 為催化主力的蛋白脢很多,統稱為 serine 型蛋白脢;除了 chymotrypsin 外,尚有許多如 trypsin (胰蛋白脢)、elastase (彈性蛋白脢)。它們的催化機制相似,整體構形相當類似,且都有 [Ser-His-Asp] 接力形式的催化機構。但對基質的專一性不同,trypsin 的基質為鹼性胺基酸 (Lys, Arg),elastase 則只切 R 基團較小且不帶電荷者。
|
|||||
|
5.4 酵素的專一性: 酵素只與特定的基質結合,有很高的專一性,這是酵素重要特性之一。 5.4.1 專一性結合區: 大部份酵素的活性區即有專一性辨識與結合基質能力,但CT另有一基質的辨識區位在其活性區附近。 a. 上述各 Ser 型蛋白脢的活性區除了有 [Ser-His-Asp] 催化中心外,附近還有一個袋狀的基質結合區,可以辨別基質的種類 (驗明正身),以便與正確的胺基酸結合。 b. 這種專一性結合是靠結合區上胺基酸 R 基團的形狀、大小、極性或電荷等性質的契合,以二級鍵結合。例如 chymotrypsin 的結合區多含非極性的胺基酸,故只能與非極性的芳香族胺基酸結合,催化這類胺基酸的水解。改變結合區的重要胺基酸,可能改變專一性 (Science, 1992, 255: 1249)。 5.4.2 專一性結合力量: 有兩大類構成因素,一是形狀的互補,另一是吸引力量。 (參考 蛋白質 2.3) a. 分子間構形互補:如『鎖頭與鑰匙 lock & key』在形狀的契合;而蛋白質分子有彈性,與基質結合可誘生更契合的構形,稱為 induced fit。因構形互補而吸引的主要力量來自凡得瓦爾力,如圖 5 中 A 與 B 之間的契合。 b. 酵素與基質間會產生吸引力量,是由兩者間的若干二級鍵所共同組成。
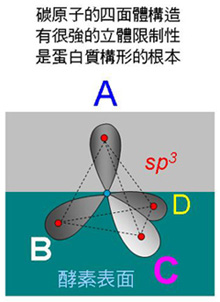
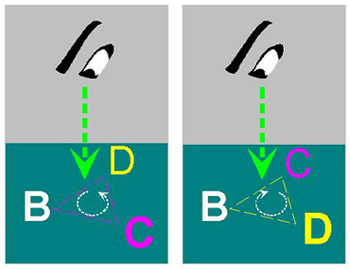
圖 5 兩分子間構形互補 5.4.3 立體專一性: 酵素分子如何辨認立體異構物? 碳的原子軌道是根本原因。 (參考 細胞與分子 圖 4) a. 酵素可辨認基質的立體不對稱性,只能對某一種立體異構物有催化反應 (如 L 或 D 型胺基酸之一),而生成物也只有對應的立體異構物之一。 b. 如圖 6 把基質分子中不對稱碳 (sp3) 四個鍵結中的一點 (A) 固定,再以酵素的專一性結合面 (深綠色) 接觸並確認 B-C-D 三點,就能分辨另一異構物的差異 (例如 B-D-C) 而排斥之 (從 A 點往下看即可分出差異,如圖 7)。
■■■ 增補幾個酵素催化機制的實例 下載連結:[Enz(5A) 2008.PPT] |
|
||||
問題集 以下包含酵素部份的全部問題。題目不一定有標準答案,甚至會引起很大的爭議,但這就是問題集之目的。1. Carboxypeptidase A 分子的活性區需要一個 Zn 離子,請問此金屬離子有何作用? 2. 酵素活性區的構形與下列三者何者較能互補?請說明為何? 基質 生成物 過渡狀態 3. 為何維持正確的酸鹼度 pH,對某些酵素的活性非常重要?有無酵素不受 pH 影響? 4. 抗體與其抗原極類似酵素及基質,有很強的專一性,但並無酵素的催化作用;而 catalytic antibody (或 abzyme) 為特別設計產生的抗體,可使抗體具有催化的性質。若有反應如︰A+B → A-B,而你要設計能催化此反應的抗體 (A 與 B 均為小分子)。 a) 你將以何種分子為抗原?詳細說明原因。 (hint: 為何酵素具有催化能力?) b) 在進行免疫時,應當注意那些事項? 5. 為何 chymotrypsinogen 分子要先經過裂解後,才會變成活性型? 6. Chymotrypsin 如何穩定所催化基質的中間過渡狀態? 7. Chymotrypsin 的 His57 對其催化機制有何貢獻?His57 如何受到環境 pH 的影響? 8. 已知 DPF (diisopropyl phosphofluoridate) 是 Ser protease 的專一性抑制劑,如何證明 DPF 是攻擊酵素的活性區? 9. 大部份 proteases 似乎都有相類似的水解機制,請說明此一核心機制。 10.胜肽鏈上可被 trypsin 專一性水解的胺基酸有哪兩種? 11.請判斷下列各題的真偽: (並說明原因) a) 水溶性蛋白質的疏水性胺基酸大都在分子的裡面,故酵素分子內部不可有親水性胺基酸,以防止分子變性。 b) 只要含有二十種基本胺基酸基團,蛋白質就可以完全發揮酵素的催化機制。 c) Carboxypeptidase A 是一種外切脢,可切除蛋白質 C-端的非極性胺基酸,它可以與 a-COOH 結合來辨識是否為 C-端。 d) Ser proteases 都有由 Asp-His-Ser 組成的 triad,可產生一具高反應性的 -O- 基。 e) 酵素催化反應進行的過程中,酵素與基質或產物之間,都不能有共價鍵生成,這才能稱為催化。 f) 蛋白脢催化過程中,所形成過渡狀態有過多正電荷,要用氧原子上的電子中和之。 g) 剛轉譯出來的完整 chymotrypsin,其催化活性很低,是因為無法穩定過渡狀態。 h) 一般酵素的活性區與專一性辨認區都不在同一位置,以便增加專一性。 i) 兩個蛋白質間的專一性結合,其構形的互補是重要條件之一,乃因於凡得瓦爾力。 j) 以下配對都是專一性吸引力所造成:酵素-基質、酵素-輔脢、荷爾蒙-受體。 12.是非選擇題 (答案寫在□內,是→○、非→╳) a) 何者為 Ser protease 家族? □ Subtilisin □ 凝血因子 VIII □ Acetylcholinesterase □ Elastase □ Thrombin b) 何者為酵素的不可逆抑制劑? □ Penicillin □ PCMB □ DFP □ EDTA □ 磺胺藥 □ cAMP c) 改變下列那個胺基酸後會影響 chymotrypsin 的活性:□ Asp102 □ His57 □ Ser14 □ Ile16 □ C-terminal d) 可構成兩個分子間的專一性吸引力的力量有那些: □ 氫鍵 □ 疏水鍵 □ 雙硫鍵 □ 凡得瓦爾力 □ 離子鍵 □ 分子間構形互補 e) 有關立体專一性何者為真? □ 碳原子的 sp3 軌道是主因 □ 自然界中多使用 L-胺基酸 □ 單糖分子並無立体異構物 □ 酵素可分辨基質立体異構物 13.請簡單解釋以下名詞:Abzyme, Metabolomics, Ribozyme, Proteasome, Specific activity
|
|
||||
|
|