|
2009 進度報告摘要 |
|
| 09/12/29 |
5 黃 惠 敏 PCS 的結晶構造與細胞定位 |
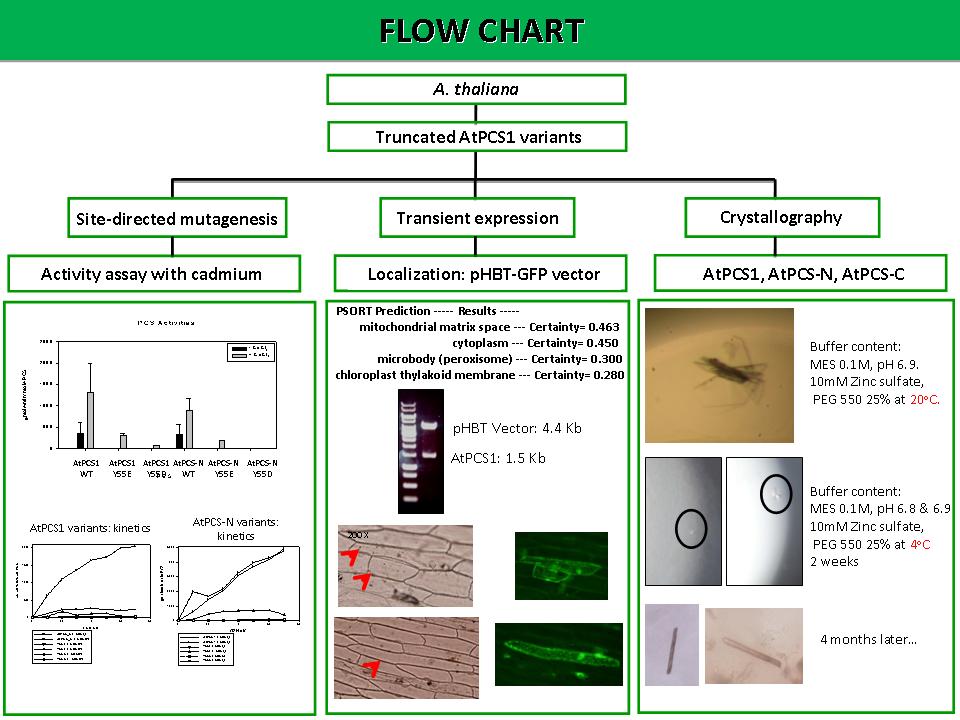 |
|
| summary |
Summary: 1) 全長 AtPCS1 加了鎘的活性最高,接著是 AtPCS1 Y55E 加鎘,再來是 AtPCS1 Y55D 加鎘。 2) 半長 AtPCS-N (加鎘與沒加鎘) 的活性沒有差異。AtPCS-N 活性最高接著是 Y55E 加鎘再來是 Y55D 加鎘。 3) 沒有加鎘的半長 AtPCS-N 突變株幾乎都沒有活性。 4) GSH substrate 濃度最高加到 30 mM。 5) Localization 預測軟體預測 AtPCS1 會在粒線體以及細胞質裡的機率較高。 6) 做了洋蔥表皮細胞的 GFP 暫時表現發現 AtPCS1 可能存在於細胞質中。 7) 利用 PSIPRED 預測 AtPCS1 二級結構發現,AtPCS1 的結構非常 flexible 使得結晶不容易形成。 8) AtPCS-N 蛋白質濃度高於 1 mg,會使得蛋白質沉澱,無法進行點晶。 Future work: 1) PCS 酵素動力學要做 Y55A。 2) 進行 substrate reaction 時必須注意每一項藥品的濃度以及加的量 3) 需了解預測軟體的原理,做 Dapi 染色標定細胞核以及打 pHBT-GFP vector 當 control 組。 4) 找做過洋蔥表皮細胞的 GFP 暫時表的學長姐討論結果。 5) 解決 AtPCS-N 蛋白質沉澱的問題。(注意 buffer 的 pH 值) 6) 繼續試出能使 AtPCS1 結晶時間縮短以及提高產量的條件。 |
| 09/12/22 |
4 謝 瑩 貞 D-enzyme 與甘藷塊根澱粉充實的關係 |
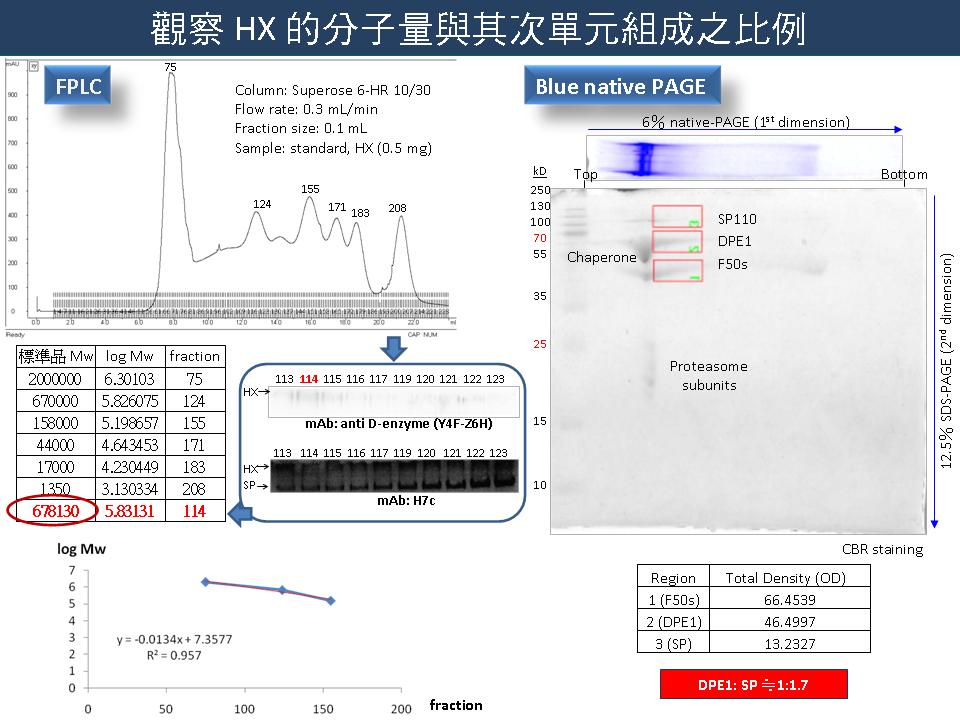 |
|
| summary |
1) Pull down assay 及 far Western 的結果並不是很清楚,應改進實驗條件,並檢討哪些步驟可能發生錯誤。 2) Pull down assay 的結果可能受到蛋白質後修飾影響,因此可先忽略表現蛋白質的部分。 3) 樣品製備是實驗的關鍵,要控制好最重要的部分,也要注意電泳過程中蛋白質量不可過多,否則會影響實驗結果。 4) 以 FPLC 觀察 HX 次單元組成之比例時,應確定 Western 所選取的分劃是否正確。 5) 通常 gel filtration 定分子量的結果不大準確,只能做為參考。 6) Blue native PAGE 應該補上 Western 結果,以確定 SP 及 DPE1 之位置。 7) 若二維電泳的蛋白質量不夠,則可能影響定量結果。 8) 利用離子交換法純化 SP 時,若以連續梯度分離蛋白質,效果可能較階段梯度好。 9) 以酵素動力學觀察 SP、DPE1、HX 間的差異時,可利用加入不同的基質量。 10) 每次實驗後都要自我檢討,並做適度的改進。 |
| 09/12/15 |
4 劉 怡 君 禽流感病毒中 RNA polymerase 的抗體製備與交互蛋白質 |
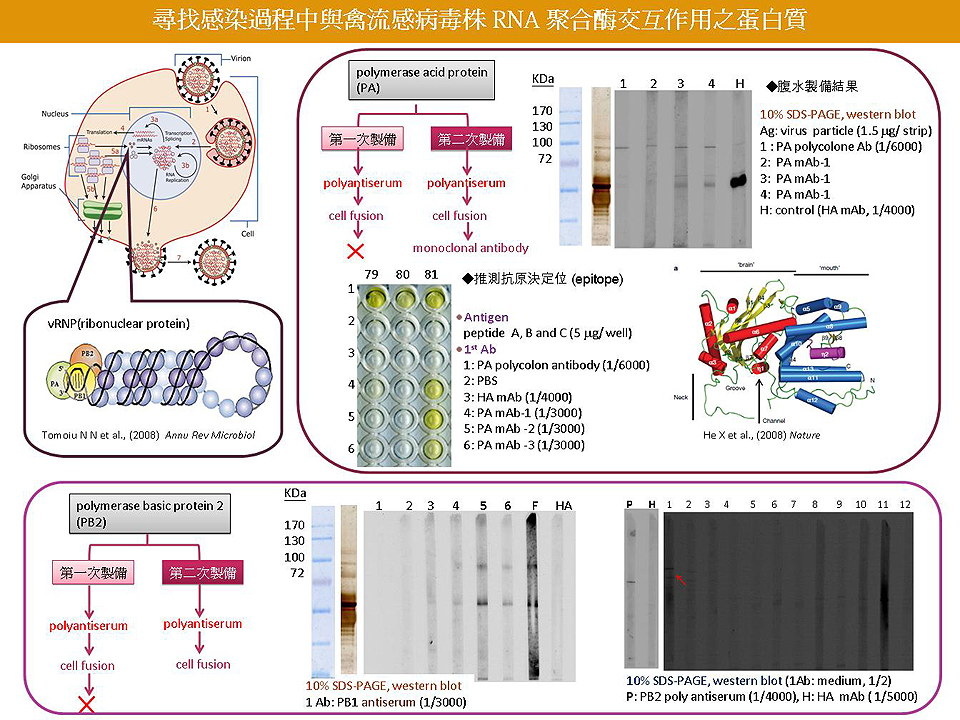 |
|
| summary |
1) 抗體製備情形:PA 已取得單株抗體,進行腹水誘導大量製備,並初步確定其抗原決定基。PB1 及 PB2 只有抗血清,需盡快得到單株抗體。 2) Fusion rate 不佳,需注意各細節,確實檢討可能的問題並改進。 3) 經腹水製備的單株抗體濃度很高,若想要用競爭的方式確認抗原決定基,必須先試出最佳的作用條件與濃度,可用等倍稀釋的方式,多做幾種稀釋倍數的測試。 4) 多株抗體的 Western blot 有非專一性結合的條帶,需改善實驗步驟,找到最佳條件。 5) 使用 PA、PB1及PB2 的多株抗體,與經雞胚培養的病毒濃縮液進行初步免疫共沉澱,需注意病毒的蛋白是否有被打破而分離開來,可參考其它論文打破病毒的方式。 6) 實驗步驟需好好設計,考慮各種情況 (設計好控制組) 以求實驗完整。 7) 工具做好了,需要應用在科學探索上,並說出完整的故事。 Future work 1) 儘快完成 PB1 和 PB2 單株抗體製備。 2) 確認現有單株抗體之 epitope。 3) 運用軟體先分析 epitope 於立體結構上之位置,或預測其他後修飾作用。 4) 推測抗體之 epitope 對病毒之功能。 5) 確認 2D-PAGE 上 PA, PB2 和 PB1 subunit 分佈。 6) 運用現有抗體以 IP 方式初步分析 polymerase 各 subunits 間的 interaction。 |
| 09/12/08 |
4 黃 迺 茵 主題 |
|
slide |
|
| summary |
|
| 09/11/24 |
1 林 歆 祐 PCS 的催化機制 |
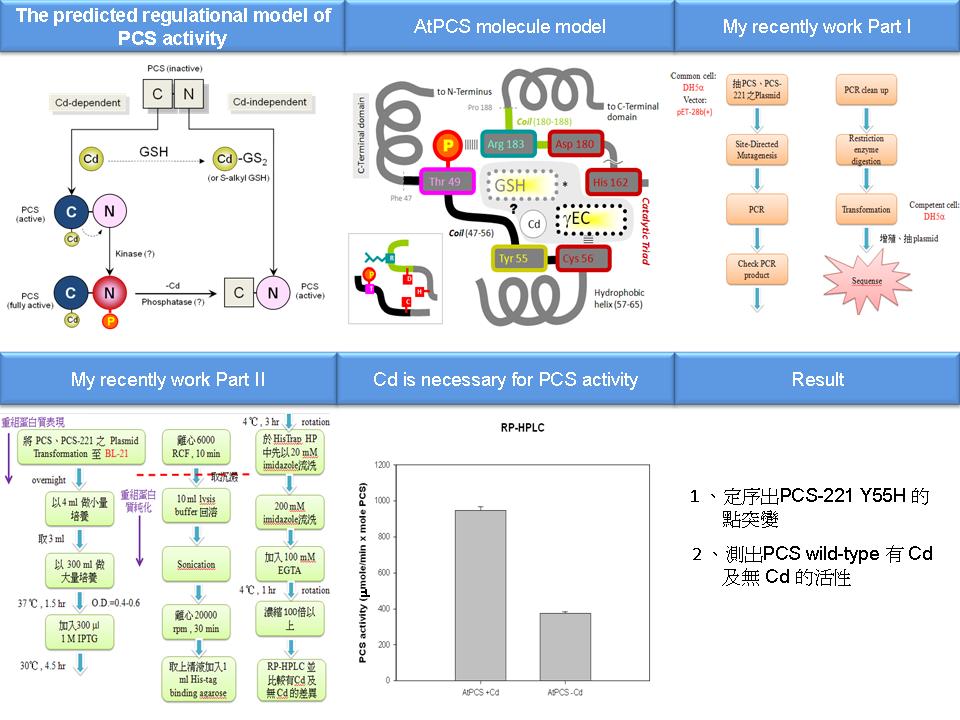 |
|
| summary |
1) 標點符號、字型格式要注意。 2) 要練習 present 說話技巧,及注意英文發音。 3) 要趕快測出 mutant 的活性分析。 4) 要把 journal 列出全名。 5) 練習說明 AtPCS molecular model 並把細節都弄清楚。 6) HPLC 後產物的計算方式是用積分值來算活性。 7) 由於 Cys 56 和 gEC 結合,可以試著把 AtPCS C-端的 Cys mutated 掉。 |
| 09/11/17 |
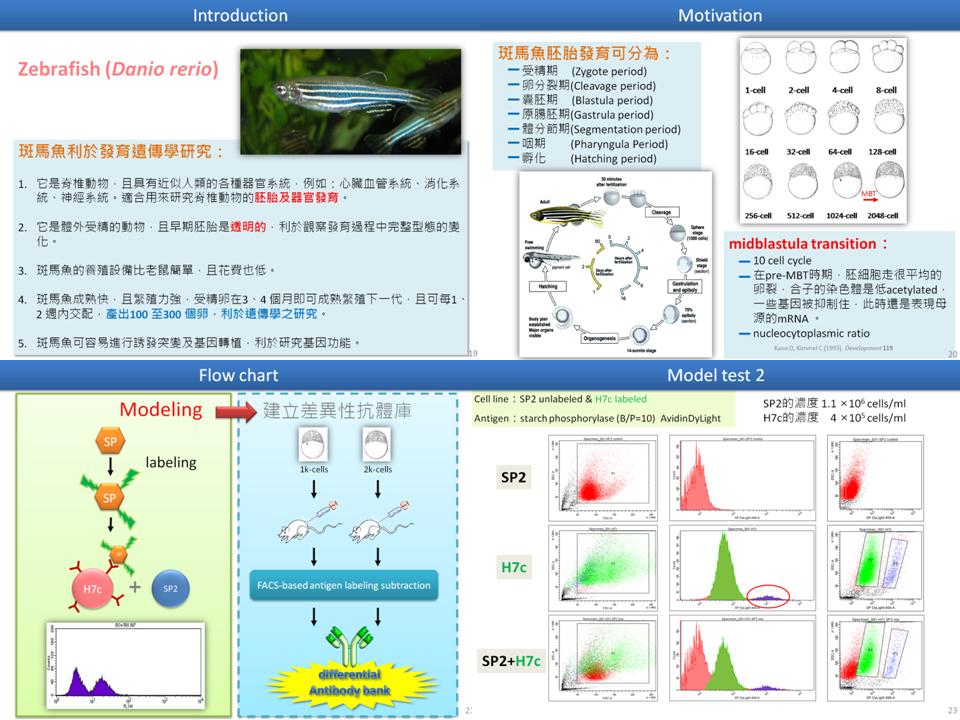
1 錢 思 翰 以流式細胞儀篩選差異性抗體庫 |
 |
|
| summary |
1) Total proteins 純化過程,可使用硫酸銨分成數個分劃下去免疫。 2) 為了使胚胎不再分裂,可以先放在冰上中止,但還是會有短暫的分裂。 3) 使用疊瓦式的免疫打法,把 sample 集中起來,分多次打,三個月打完。 4) 剛 fusion 的 hybridoma 也許效價會不穩定,也許可以在 fusion 後養久一點? 三週或四週後再篩選? 5) 為何使用 1K 或 2K 細胞? 做母源轉胚源應該用 1K 或 2K? 6) 一個 cell cycle 需要 15 分鐘,先做了 deyolking,再跑 2D 後,再選擇使用哪個時期。 7) 512~1024 是第 10 個 cell cycle,不過為了找出較大的差異,可以選擇 128 和 2K 做比較。 8) 先用流式細胞儀做一次全流程,不行再試磁珠。 9) 準備多隻老鼠免疫,以備接續實驗不斷。 10) 為何合成的 peptide 分子量那麼大? 11) GST tag? (只寫這樣看不懂) |
| 09/11/10 |
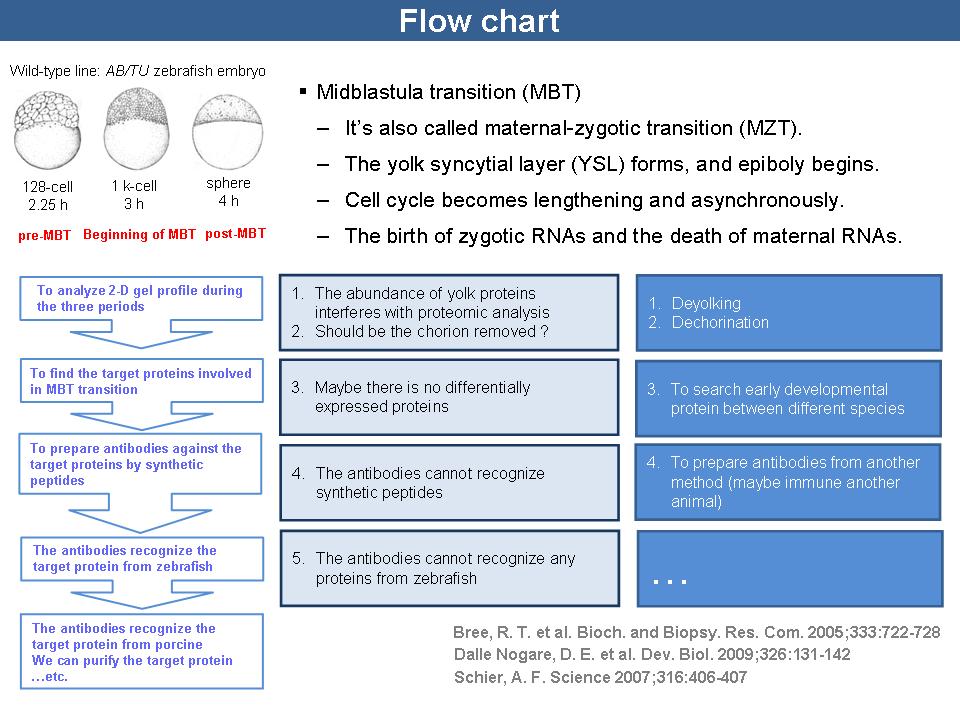
1 施 驊 珊 斑馬魚卵發育過程的蛋白質體表現比較 |
 |
|
| summary |
1) Lucitt 等人於 2008 發表的論文中,針對斑馬魚胚胎 70 hpf 及 120 hpf 時期,利用 2-DE 電泳及 LC-MS/MS 進行蛋白質體研究,有無後續研究? 2) 研究 MBT 是否真的只限於基因方面的研究,蛋白質體方面的研究是否真的大都針對發育後期? 3) 細胞數越少的時期,蛋白質量是否越少? 4) 斑馬魚胚胎的 MBT 從 512-cell 開始,許多文獻指出 128-cell 時期為 pre-MBT,sphere 時期為 post-MBT。但由於 128-cell 和 512-cell 太過接近,故改選擇 128-cell、1 k-cell 及 sphere 三個時期進行 2-DE 電泳。 5) 2-DE 電泳的樣本前處理,需經去除卵殼 (dechorionation) 及去除卵黃 (deyolking) 等步驟以減少干擾。而經 dechorionation 後的樣本,進行 deyolking 效果較佳。 6) 利用低電壓 (75 V) 進行 SDS-PAGE 使得蛋白質點擴散嚴重,故還是以 120 V 進行第二維電泳。 7) 根據 Link 等人發表的文獻中,樣本處理的 IEF program 電壓達到 10,000 V,可能是造成蛋白質點擴散的原因,建議可設為 8,000 V 達到相同 Vhr 來進行 IEF。 |
| 09/10/20 |
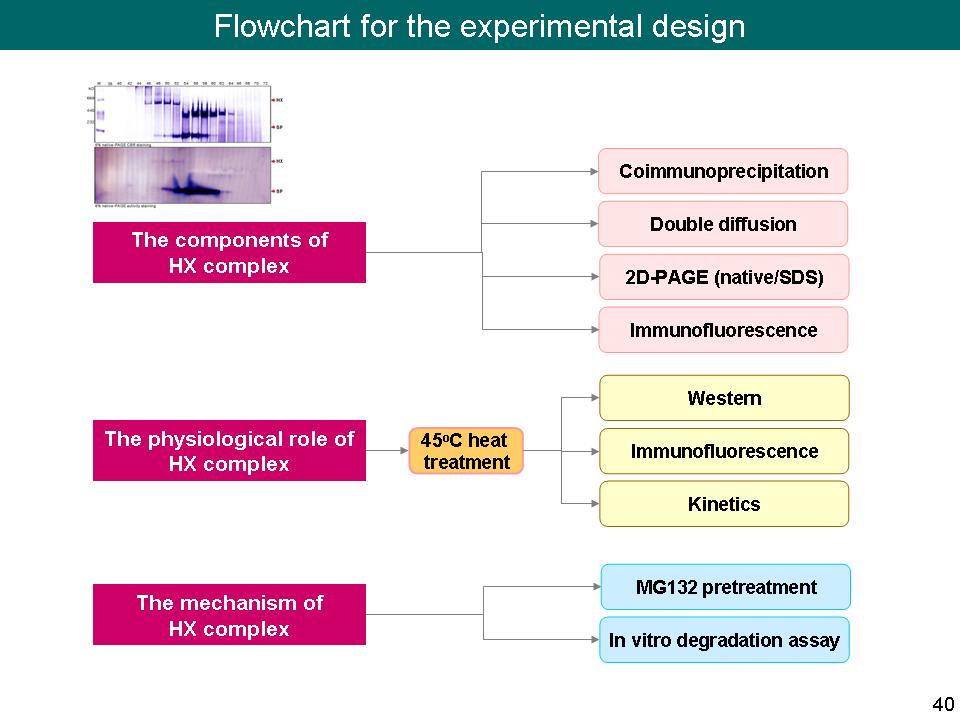
10 林 怡 岑 L-SP 分子上 L78 之降解受到溫度與 proteasome 的控制 |
 |
|
| summary |
實驗結果: 將甘藷塊根切片進行 45ºC 加熱處理後,隨著時間累積,L-SP 有階段性降解的現象,目前推測 first cutting site 在 PEST motif 上,並由 proteasome 機制降解 L78,此結果可能改變 L-SP 的生理活性,由合成澱粉轉為磷解澱粉,產生 Glc-1-P,進而促進 NADPH 生成,來減少因熱逆境產生 ROS 所造成的細胞傷害。 建議事項: 1) 問問看張老師有沒有 19S regulator 之 antibody,利用 19S regulator antibody 進行 Western,觀察 19S regulator 是否存在,雖然之前利用 LC/MS/MS 都沒有發現,而且目前也發現一些 proteasome 降解機制不需要 19S regulator 參與。 2) 將 45ºC 加熱處理不同時間之 L-SP 降解片段進行 N-端定序,看看斷裂的機制是否符合 Belizario et al. Curr Protein Pept Sci. (2008) 9(3): 210-220 所提出的,先由 caspase 切一刀後,再經由 20S proteasome 降解之機制。 3) Caspase 專一性切位是什麼?主要是切 Asp 的 C-端;而 Belizario et al., 2008 則提出 caspase 會辨認 PEST motif 並降解之。 4) 植物中含有 caspase 嗎?它與細胞凋亡及其他很多調控作用很有關係,查看植物中 caspase 的相關研究。 5) Confocal images 都是用同一條件去照射的嗎? 是的,相同強度,相同時間。 6) 目前推測整個機制的 trigger 是 heat shock,L-SP 受熱開始經由 proteasome 路徑降解,cutting site 在 PEST 上,L78 去除後,L-SP 傾向磷解澱粉產生 Glc-1-P,然後呼應 Zeeman et al. Plant Physiology (2004) 135: 849-858 所提出,Glc-1-P 可能進而促進 NADPH 生成,來減少 ROS 所造成的細胞傷害。如果真是如此,我們可以追蹤這條路徑? 7) 看看陳翰民博士的論文中,L-SP 的切點為何?能不能符合 caspase 的切位? 8) 加熱到 32 h 後切片還容易嗎? 有沒有糊化?Confocal images 要附相對之光學顯微鏡 images。 9) 查閱 gelatinization 和 heat shock 有沒有關係? Gelatinization 是我們很早就觀察到的現象,可以從中查到更多的文獻,看看植物中澱粉 gelatinization 代表什麼? 10) Kinetics 作了幾次? 要記得加 error bar。 L-SP 磷解方向測 Glc-1-P 生成,反應好不好作?要用耦合反應。 11) MG132 看來的確可以抑制 L-SP 降解,但無法觀察到 L-SP 有 ubiquitination 現象。 12) 有沒有什麼方法可以證明 L78 進入 20S proteasome (DM) 的活性口袋? 13) 45ºC 加熱處理可以再做久一點,看看 L-SP 到最後會如何? 14) 生理情況下會發生連續 24 h 以上 45ºC 的高溫嗎? 我們的實驗算是放大整個逆境的訊號,來觀察整個 pathway,在生理上,可能不需要那麼長的時間,在甘藷塊根表面上的一些細胞就開始發生這些作用。 未來工作: 1) 利用 N-端定序找出 45ºC 加熱處理後,L-SP 的第一個斷裂點。 2) 將 L78 上面的幾個斷裂點胺基酸進行 point mutation,看看降解現象是否會有差異。 3) 另外,可買 caspase 來試試,看能不能切 L-SP 的 PEST motif,然後搭配純化 20S proteasome,模擬整個降解機制。 4) Heat shock, caspase, proteasome, L-SP 以及 gelatinization,找出它們之間的關連性。 |
| 09/10/13 |
6 賈 儒 珍 主題 |
|
slide
|
|
| summary |
|
| 09/09/29 |
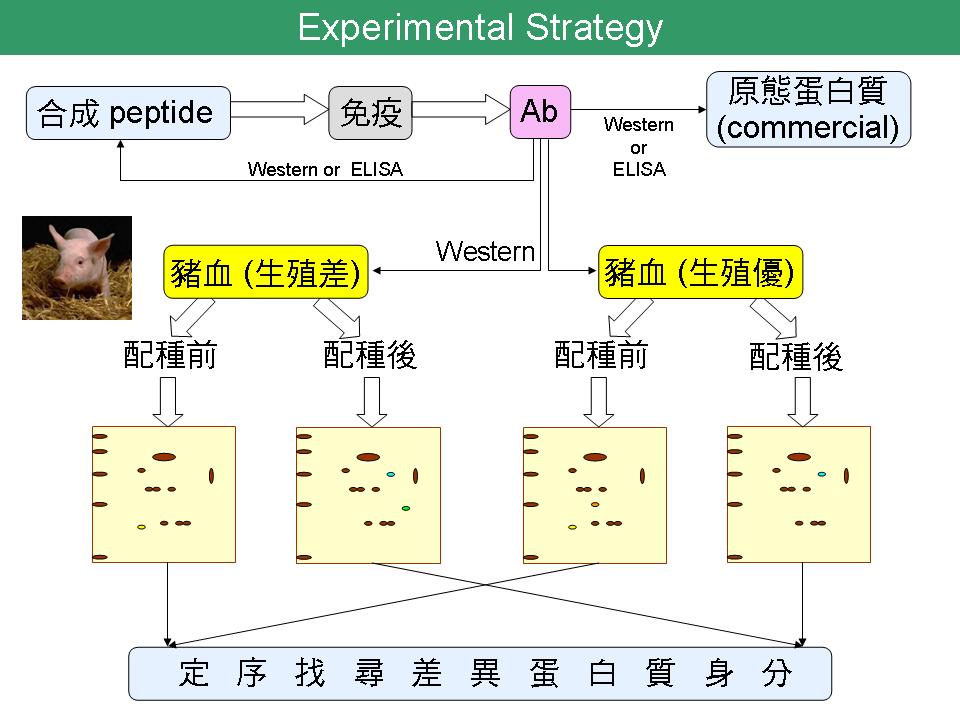
1 沈 志 昱 母源轉換到胚源的標誌蛋白質及其抗體製備 |
 |
|
| summary |
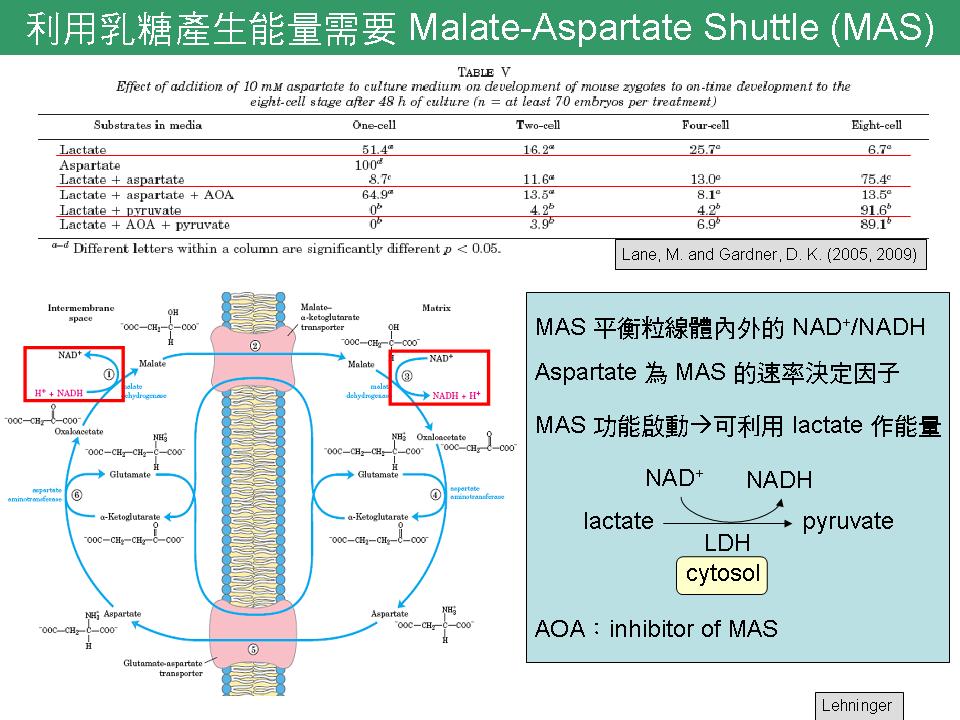
Part 1 1) 合成 peptide 挑選序列時,盡量避開 a helix。此外,轉譯後修飾也須考慮在內。 2) 引用文獻時,須加註期刊出處。 3) IGF-1 與 IGFBP3 的關係: 培養基中添加 IGF-1 能增加發育至囊胚期的胚胎比例,大部分 IGF-1 會與 IGFBP 結合,其中以 IGFBP3 為最主要蛋白質。IGFBP 與 IGF-1 結合,能增加 IGF-1 的半衰期,換句話說,IGF-1 的作用時間將延長。此外,與 IGFBP 結合能調控其運輸與位置。雖然 IGFBP 在早期胚胎發育的角色尚不清楚,但已有研究指出 IGF-1 會改變 IGFBP 在胚胎中的表現量,而 IGFBP 能控制 IGF-1 的反應。 4) 以 peptide 製作抗體時,可能遇到下列問題: (a) 缺少原態蛋白質,無法確認抗體的真實性。 (b) 就算抗體能辨認原態,若這三種蛋白質在豬血液中含量太少,也無法偵測。 → 搜尋此三種蛋白質有無在人類的資訊,再來評估。 (c) 承上,可將抗體接在膠體上,利用親和層析管柱濃縮該蛋白質後再行偵測。 Part 2 Summary slide 1) Lane & Gardner 在 2005 年於 JBC 研究指出,必須啟動 malate-aspartate shuttle (MAS) 的功能,胚胎才能利用 lactate 產生能量。因為 lactate 必須先在 LDH 作用下轉換成 pyruvate,故認為 MAS 可能與 LDH 有 coupling 且互相影響。 → 除了追蹤 Lane & Gardner 的研究外,擴大搜尋有無與能量代謝相關的證據。 |
| 09/09/22 |
3 謝 瑩 貞 D-enzyme 與甘藷塊根澱粉充實的關係 |
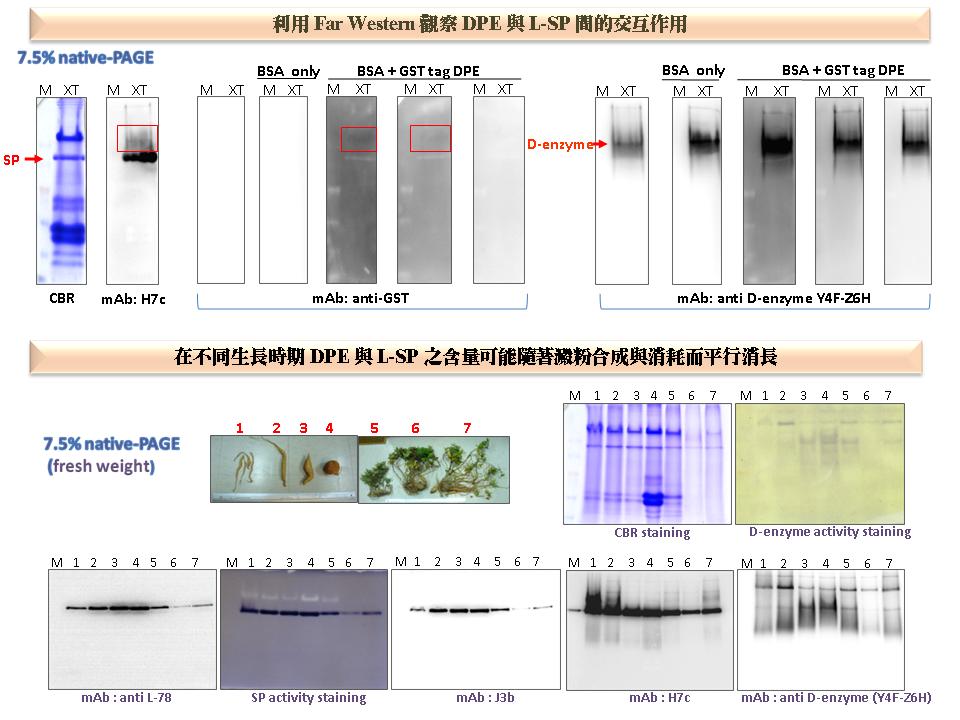 |
|
| summary |
1) 由於 Far Western 結果不是非常明確,應改進實驗條件以提升訊號強度,或利用其他方法來觀察 L-SP 與 D-enzyme 之作用情形。 2) 評估是否可利用綠藻 (515) 研究 L-SP 與 D-enzyme 間之交互作用或生理意義。 3) 利用相同來源,不同大小的甘藷塊根以表示各階段的澱粉充實程度,觀察其中 L-SP 與 D-enzyme 之含量,並進一步探討蛋白質在不同生長階段的重要性。 4) 進行原態電泳時,可延長電泳的時間,以利高分子量的蛋白質分開。 5) 實驗前應先想好研究動機,並加強實驗結果的呈現。講者必須站在聽眾的角度,以說故事的方式描述實驗目的及結果之含意。標題應明確顯示該實驗之意義與重點,使聽眾能一目了然。 6) 應設計其他實驗方法,進一步探討 D-enzyme 與 L-SP 之作用,在植物生理上是否有重要的影響?多看論文多與別人討論,可以啟發對研究的新想法,有利於尋找實驗的動機。 |
| 09/08/20 |
3 劉 怡 君 RNA polymerase 在 AIV 感染過程的角色 |
 |
|
| summary |
1) 使用兩株本土禽流感病毒 (台大獸醫系王金和) A/chicken/Taiwan/2838V/00 (H6N1);A/chicken/Taiwan/2838N/00 (H6N1) 2) 目前已得到可分泌 PA 單株抗體之細胞株,將繼續放大繼代,盡速取得純化抗體。 3) PB1 及 PB2 的單株抗體仍在製備中。 4) 可先與不同時期經病毒感染的宿主細胞進行免液共沉澱 (IP),確認抗體確實可抓到宿主細胞株內的病毒 RNA 聚合酶,並儘快取得初步結果。 5) 比較不同時期經病毒感染的宿主細胞 2-DE 圖譜上蛋白質點是否有差異,若存在差異則以 mass 鑑定身分。 6) 定義『不同感染時期』,參考文獻看看是否有 maker protein 或 marker gene。 7) 確認抗體染色的位置是否具有特殊意義。 8) 使用共軛焦顯微鏡與免疫螢光染色,直接觀察三種 RNA 聚合酶於宿主細胞內組裝的過程。 |
| 09/07/15 |
3 黃 迺 茵 阿拉伯芥的 PCS 突變株及點突變 PCS 構築 |
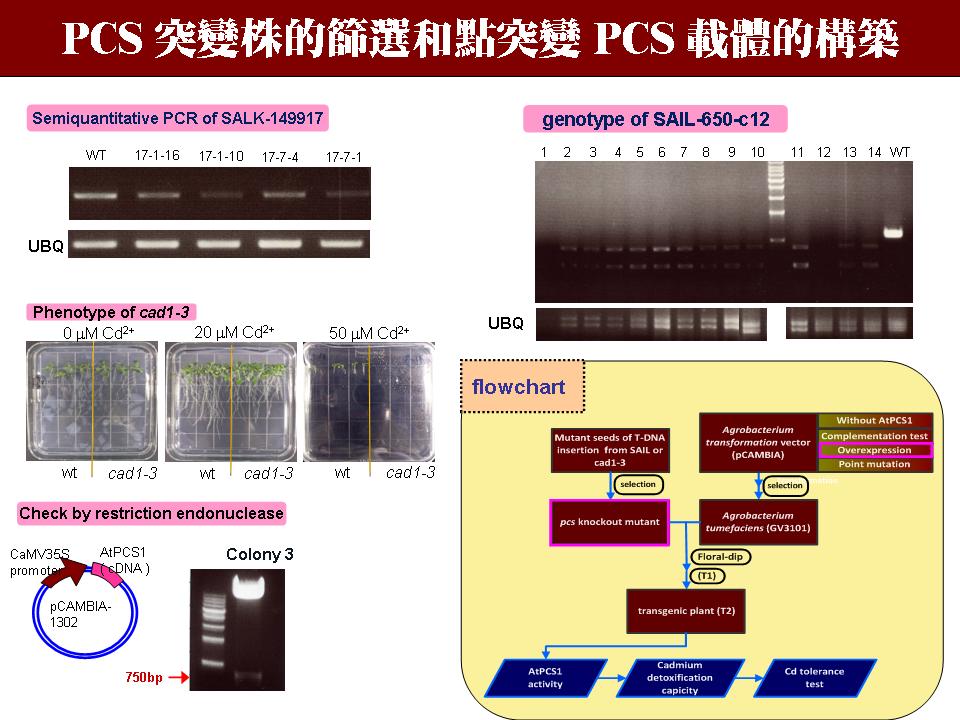 |
|
| summary |
1) 由於 DNA 最後的產物是蛋白質,所以也可以直接用抗體去偵測 PCS 的表現量,但是阿拉伯芥中的 PCS 量是否足以被抗體抓到,且專一性如何,這需要進一步實驗證實。 2) 雖然 Cad1-3 是已經發表的突變株,還是要再次確認它的酵素活性和性狀,最好可以重現 paper 內的性狀。 3) 要注意實驗進度,最好連點突變株的載體一起構築,自己要抓緊時間 (2009/12/31 的目標?)。 4) 由於植物轉殖等待收種子的時間很長,這期間可以進行其他實驗,例如測試點突變表現蛋白質的性狀,或是純化內生性的 PCS 等。若 in vitro 的性狀跟預期不符合,則無法做到 in vivo。 |
| 09/07/15 |
5 黃 惠 敏 PCS 的結晶構造與細胞定位 |
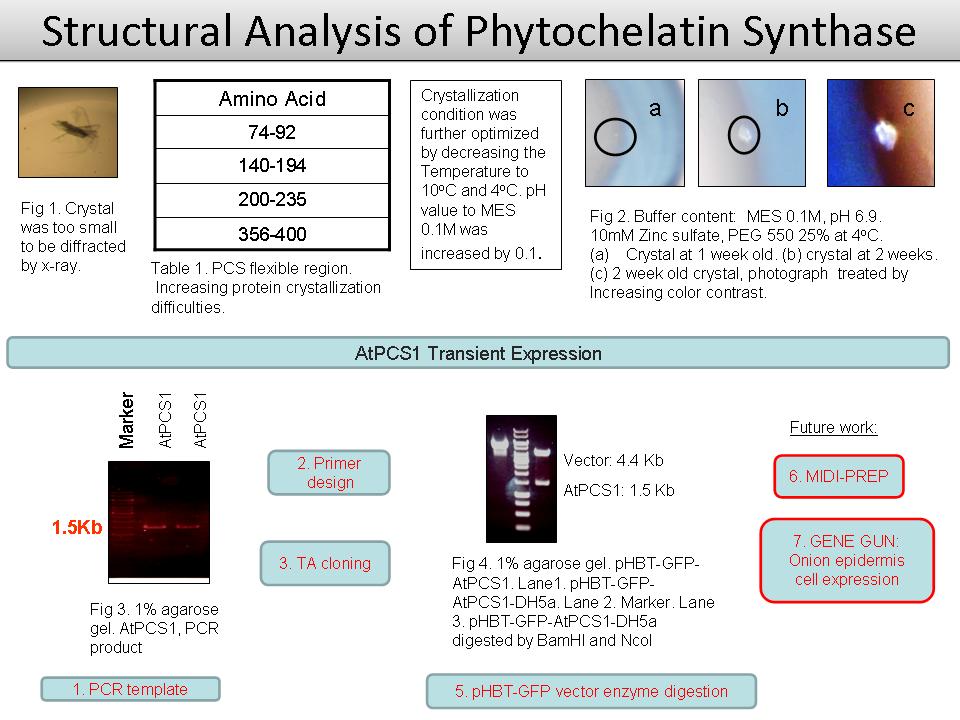 |
|
| summary |
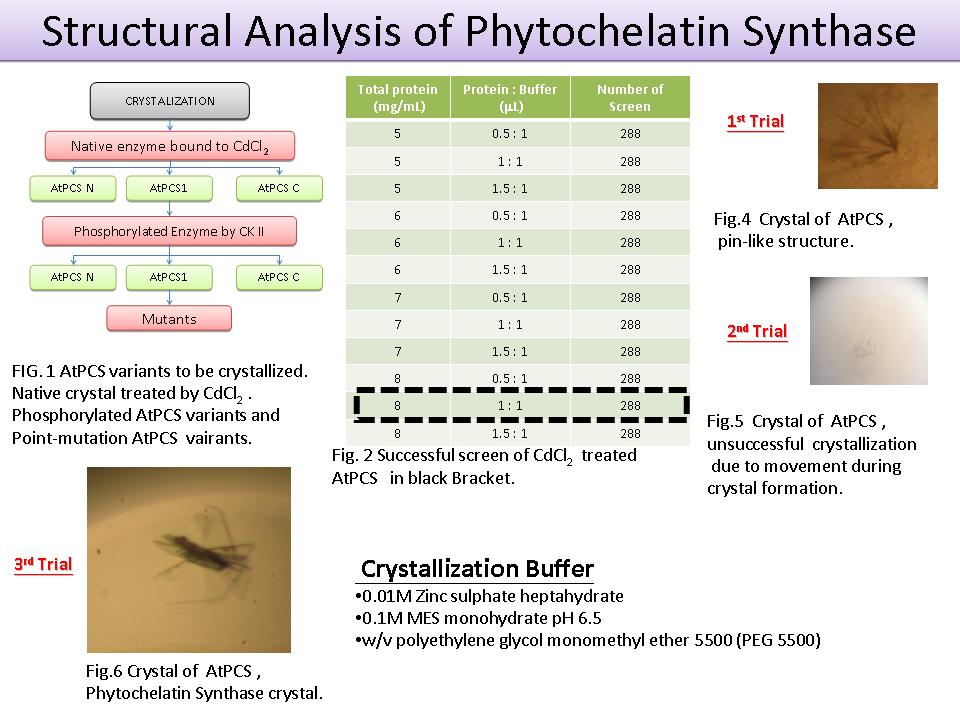
1) 目前所結出來的晶體因太小無法進行 X-ray 繞射。 2) 結晶 buffer 的 pH 值以 0.1 進行微調。 3) 使用不同溫度:25ºC / 10ºC / 4ºC 4) 微調後晶體 buffer:0.1 M Zinc sulphate heptahydrate, 25% w/v PEG 550, 0.1 M MES, pH 6.9 (4ºC)。 5) 晶體由三個月的生長時間縮短至兩週。 6) 因 cold room 震動太大,會影響晶體的結構以及形成時間。 7) 使用 PSIPRED (UCL) 模擬 PCS 二級結構發現:PCS 結構有許多 flexible area,因此造成晶體形成困難。 8) PCS 進行 GFP 細胞定位實驗:探討 PC 合成的位置,了解重金屬在進入細胞後能被捕捉的位置。 9) 已的完成 pHBT-PCS-GFP 的 cloning。 Future Work: 1) 結晶找到更快速生長條件,未來需要進一步找出使結晶變大的條件。 2) 進行 pHBT-PCS-GFP 的洋蔥表皮細胞內表現。 |
| 09/05/27 |
Y 羅 庭 芳 禽流感病毒 H1 isoforms 上面的醣化現象 |
 |
|
|
summary |
1) Comparisons of the 2-DE patterns for 2838N and 2838V showed major differences in hemagglutinin 1 (HA1). 2) HA1 contained six isoforms with similar molecular mass but showing different isoelectric points (pI). 3) The difference in pI values was not due to the phosphorylation on HA1. 4) MALDI MS and MS/MS analyses for the N-glycans of the HA1 isoforms revealed that sialic acid was not included. 5) The isoform with lower pI showed higher percentage of an unknown glycan with m/z at 1867 (X-glycan). 6) These HA1 pI isoforms showing similar molecular mass but distinct pI might due to the effect of X-glycan on the molecule. 8) It is possible that the special glycosylation on HA1 might have effects on host recognition, as well as tissue tropism or immune escape. |
| 09/05/20 |
9 林 怡 岑 L78 影響 L-SP 之降解與催化活性 |
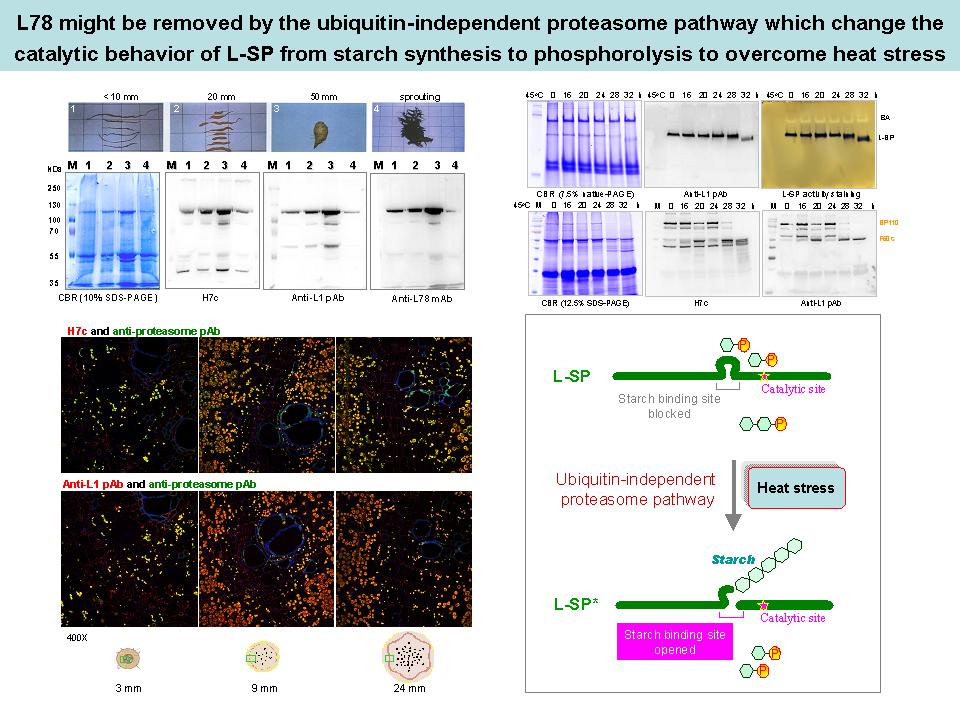 |
|
| summary |
實驗結果: 1) L-SP 在不同甘藷塊根生長時期,其分子上的 L78 並不會斷裂,但是將甘藷塊根切片進行 45°C 加熱處理後,發現隨著時間的累積,L-SP 有階段性降解的現象,此結果顯示 proteasome 所造成的 L78 降解,可能改變 L-SP 的生理催化活性,由合成澱粉轉為磷解澱粉,產生 Glc-1-P 來抵抗熱逆境。 2) L-SP 與 D-enzyme 之蛋白質含量都隨著甘藷塊根的生長而增加,並且在成熟甘藷塊根中,兩者互相結合的現象最為明顯,此結果顯示這兩種蛋白質可能在成熟甘藷塊根中,以 protein complex 的形式進行 MOS 的回收與重新利用。 建議事項: 1) 阿拉伯芥葉片並不適合用來觀察澱粉是否累積之表型,因為其為 source tissue,所累積的是暫存性澱粉。 2) Satoh et al. The Plant Cell (2008) 20: 1833-1849 中,是如何推論 L-SP 與 primer initiation 有關? 3) 加熱處理之甘藷塊根可進行冷凍切片之 confocal microscopy,另外,加熱時間可延長至 36 hr。 4) 觀察甘藷塊根不同生長時期之 L-SP 含量變化,可分為固定蛋白質總量與固定 fresh weight 兩種方式來比較。 5) Review Satoh et al. The Plant Cell (2008) 20: 1833-1849 與 Tetlow et al. Plant Physiology (2008) 146: 1878-1891 之文獻。 6) 利用生物資訊學比較不同物種 L-SP 之序列 (尤其是會累積澱粉的物種),可由這些資訊獲得一些 information and new ideas。 7) 由初步的 ubiquitination 實驗,無法斷定 L-SP 是否受到 ubiquitination 之修飾,因為 ubiquitination 之蛋白質可能很快就被降解而不易觀察,而 MG132 的抑制效果也有限。 8) 將古生菌 20S proteasome deletion mutant (DM) 造成 L-SP 斷裂的 F50s 片段進行 N 端定序,可得知 major cutting site 為何,此實驗結果可與陳翰民 (1997) 之實驗結果作比較。 9) L-SP 與 HX 對於 soluble starch 之 Km 值比較,並不具重大意義。 10) E. coli 中的 MalP 比較類似植物中的 L-SP 還是 H-SP? Ans:E. coli 有兩種 phosphorylase,分別為 MalP 和 GlgP,兩種 phosphorylase 的分子序列中均不含類似 L78 之序列。 11) 應比較 E. coli 中 MalP 和 GlgP 與馬鈴薯或其他物種中 L-SP 和 H-SP 的相似度。 12) Glucan trimming model 中有說明 maltotriose 會累積嗎? Ans:Myers et al. (2000) Recent progress toward understanding biosynthesis of the amylopectin crystal. Plant Physiol 122: 989-997 中推測 maltooligosaccharides (MOS) 會累積。 13) 陳翰民 (1997) 有比較過完整的 L-SP 與斷裂的 L-SP 對於澱粉的親和力改變,但並未比較過對於 MOS 的親和力改變,可參考此實驗,比較完整的 L-SP 和斷裂的 L-SP 對於 MOS 的親和力改變。 14) 有 paper 提出 L-SP 和 D-enzyme 會形成 protein complex 嗎? Ans:沒有,目前科學家只推測此兩種蛋白質所進行的是連續性的反應。 15) 參考 Tetlow et al. Plant Physiology (2008) 146: 1878-1891 所進行的實驗,設計關於 protein-protein interaction 之相關實驗。 16) Confocal 實驗中 L-SP 和 D-enzyme 的 merge 結果相似度太高,可調整抗體濃度或其他條件來改善。 17) Confocal 實驗中 L-SP、proteasome 和 D-enzyme 都位於 amyloplast 中,位置都很相近,有沒有可能這三種蛋白質互相結合?可進行 proteasome 和 D-enzyme 之 confocal 實驗,觀察兩者是否有 co-localization 之現象。 18) L-SP 和 proteasome 互相結合,可是 L78 卻依然存在,此現象如何解釋? 19) 論文中需詳加敘述關於 E. coli 中 malP (L-SP) 和 malQ (D-enzyme) 的關係,因為那是整個故事的起源。 20) 是否嘗試過將 L-SP 和 D-enzyme 在試管中混合? Ans:有,但結合力很弱,無法明顯觀察到。 21) 由 GST pull-down assay 的結果推測 crude extract 中,可能有 something 幫助 L-SP 和 D-enzyme 結合,或是 L-SP 受到某種後修飾調控。 22) 關於 L-SP、proteasome 和 D-enzyme 這三個蛋白質的關係,你打算怎麼解釋? Ans:希望能分別進行探討。 未來工作: 1) 未來方向需著重兩個蛋白質互相結合之生理意義,以及結合的 mechanism,例如為何需要 proteasome?proteasome 如何調控 L78 降解?以及 L-SP 與 D-enzyme 之 pathway 等。 2) 以寫 paper 的心態開始整理 data,一篇 paper 大概需要七個圖與幾張表,每張圖要能說明一個故事。 |
| 09/05/13 |
5 賈 儒 珍 主題 |
|
slide
|
|
| summary |
|
| 09/04/29 |
12 何 杰 龍 主題 |
|
slide
|
|
| summary |
|
| 09/04/08 |
2 謝 瑩 貞 SP 與 D-enzyme 的蛋白質交互作用 |
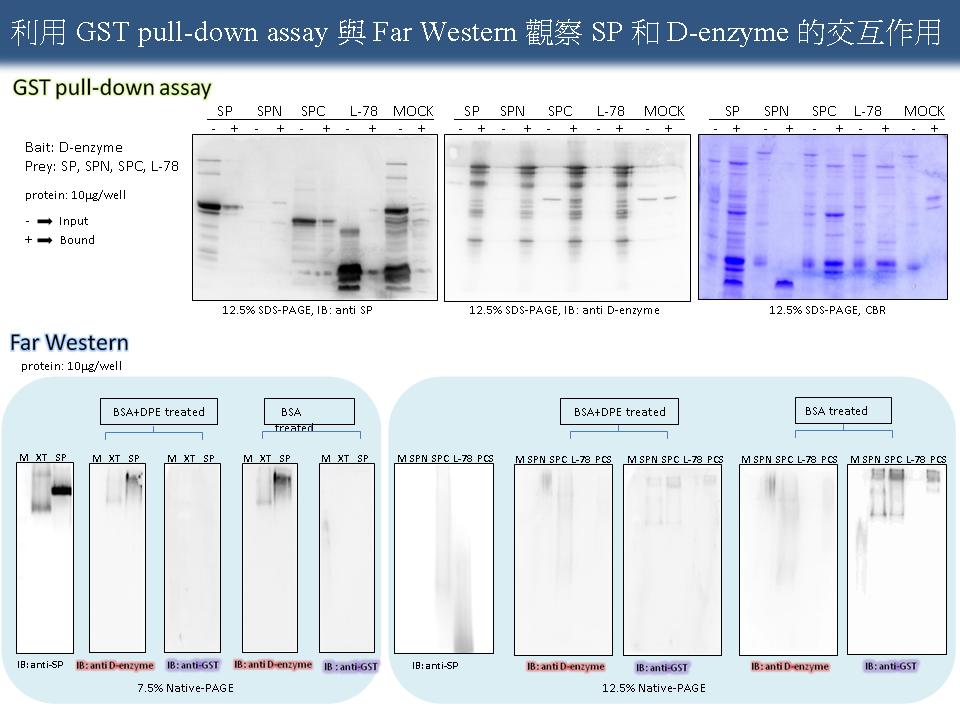 |
|
| summary |
1) SP 的粗抽液與硫酸銨一起沉澱下來,如此的蛋白質可以在 -20°C 儲存。 2) 引用別人的 slide 須標明來源。 3) 跑電泳時,每個 well 必須加入同樣的蛋白質量,且避免蛋白質過多,影響結果的可信度,每張 slide 上必須註明跑電泳時所加入的蛋白質量。 4) XT 請以『粗抽液』表示。 5) SPN 與 SPC 非全長,其構型可能無法摺疊完全,因此可能影響與 D-enzyme 的結合力。 6) 應聚焦思考蛋白質交互作用可能的生理意義,並著重於整體澱粉生理上的研究。 7) 由於 SPN 與 SPC 表現的蛋白量太少,且 pull-down assay 結果產生許多非專一性訊號,未來將改用 Far Western 觀察在植物體中 SP 和 D-enzyme 之間的關係。 8) 利用不同生長時期的 SP 粗抽液,和不同純化階段的 SP 進行 Far Western,並加上 anti D-enzyme 當對照組,以利分別內生蛋白質與 D-enzyme 表現蛋白,並控制相同的曝光時間進行 ECL 呈色 。 9) 應徹底了解 SP 與 D-enzyme 的生理功能與其參與澱粉代謝的目的。 10)可將不同 anti-SP 的單株抗體混合使用,取代傳統性血清。 11)利用 BiFC 進行 in vivo analysis。 |
| 09/03/25 |
2 劉 怡 君 製備 AIV 病毒蛋白 PA 及 PB2 單株抗體 |
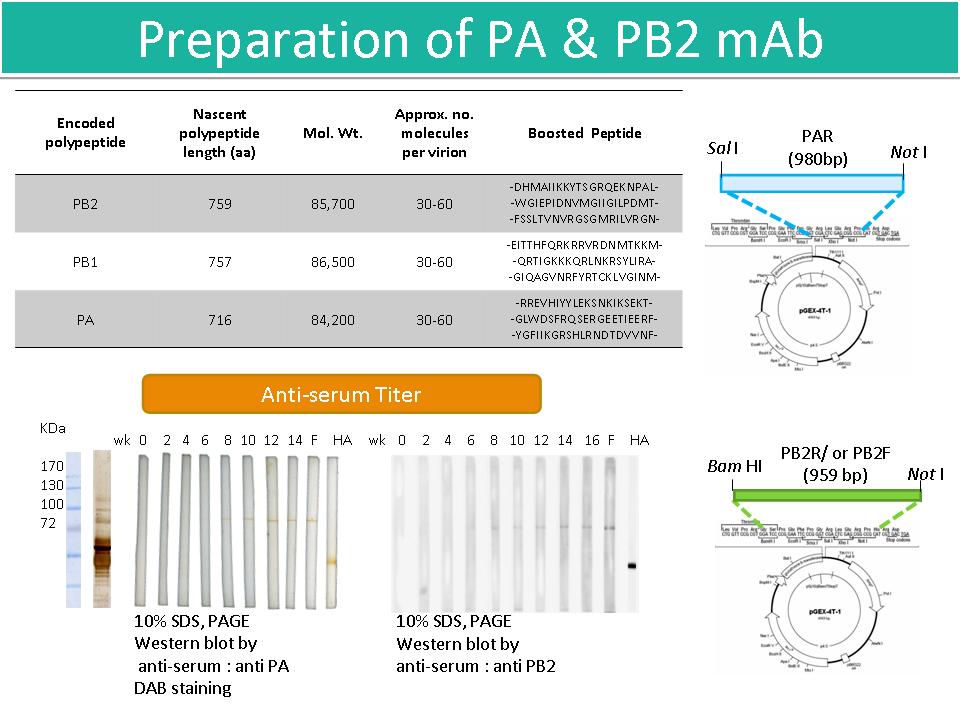 |
|
| summary |
1) 使用 2 株本土禽流感病毒 (台大獸醫系王金和老師提供) A/chicken/Taiwan/2838V/00 (H6N1);A/chicken/Taiwan/2838N/00 (H6N1)。 2) 深入了解研究 PA & PB2 的目的,製備 PA & PB2 mAb 有何作用? 以科學研究目的領導實驗方向。 3) 已將免疫後的小鼠 fusion,得到 PA及 PB2 的 anti-serum。 4) 使用 PA 及 PB2 anti-serum,嘗試定位兩種蛋白質於 2-DE 上位置。 5) 比對用來免疫的 peptide 位於蛋白質立體結構上的位置,找出其應用的生理意義與功能。 6) 利用 pGEX-4T-1 vector construct PAR, PB2R 及 PB2F;也嘗試利用 2-DE 電泳分開兩種蛋白質,希望得到獨立的蛋白質後,進行 Far Western。 7) 因為同時擁有 PA 及 PB2 的 anti-serum,因此可能朝向 binding site 或是 protein-protein interaction 的方向搜尋。 |
| 09/03/18 |
2 黃 迺 茵 PCS 在阿拉伯芥中的表現與調控 |
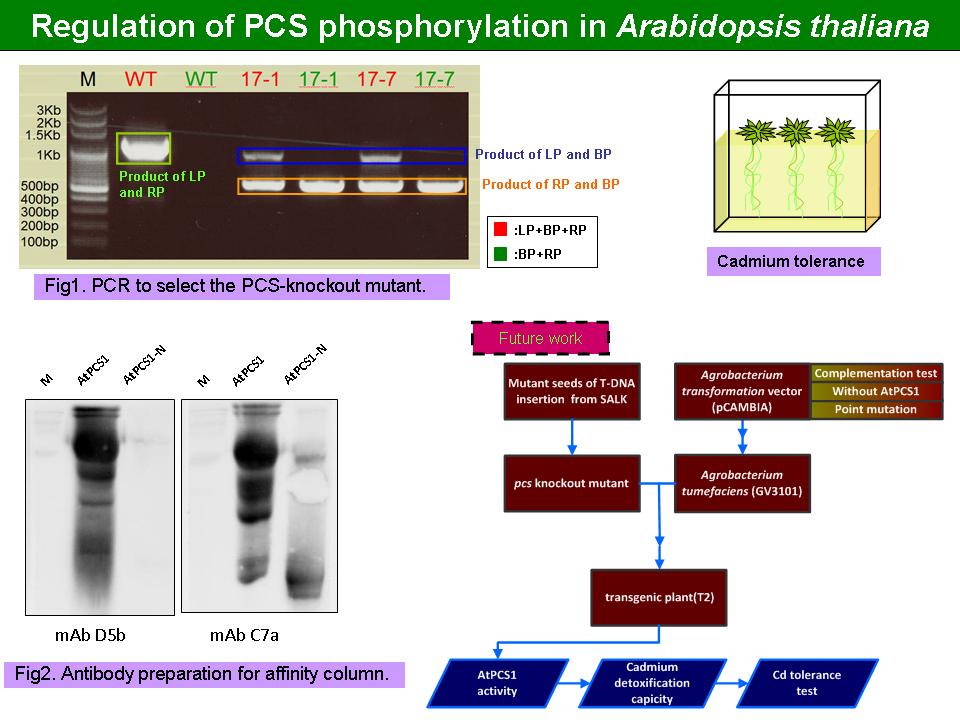 |
|
| summary |
1) 已經篩到 homozygote。有研究顯示出,PCS 基因缺失會造成阿拉伯芥在面臨鎘逆境時無法合成 PC,外觀上葉片呈現萎黃狀,且根部會有明現的褐色色素沉澱。 2) 應設計好 cadmium tolerance 的分析方法,比較的組別有哪些 (例如 PCS-null, PCS-point mutant 和 WT 在鎘處理下的根長度會因鎘耐受性之不同而有差異)。 3) 應釐清實驗的主軸是什麼 ? 目的是什麼 ? 並學習以有條理的方式表達。 |
| 09/03/11 |
4 黃 惠 敏 Phytochelatin synthase 的分子結構分析 |
|
|
|
| summary |
1) 氯化鎘處理過後的 AtPCS1 在蛋白質與結晶液體 1:1 的比例下可形成結晶。 2) Total protein 濃度 8 mg/mL 3) 結晶 buffer: 0.01 M zinc sulphate heptahydrate; 0.1 M MES monohydrate, pH 6.5; polyethylene glycol monomethyl ether 5500 (PEG 5500) Future work: 1) 增加晶體的體積。 2) 縮短結晶生長的時間。 3) 找出 AtPCS-N 與 AtPCS-C 的結晶條件。 4) 將 PCS-GFP 表現於洋蔥表皮及阿拉伯芥原生質體細胞,藉以觀察細胞中位置。 |
| 09/03/04 |
4 羅 庭 芳 AIV 上 HA1 的醣類修飾 |
 |
|
| summary |
1) 使用 2 株本土禽流感病毒 (台大獸醫系王金和老師提供) A/chicken/Taiwan/2838V/00 (H6N1);A/chicken/Taiwan/2838N/00 (H6N1) 2) 2838V 由 2838N 突變而來,HA 突變率 1.41% 共有八個突變位,兩者 HA1 peptides 相差三個 charge。 3) Lectin blot 結果直接證明 HA1 上有 a2-3 linkage 之 sialic acid。 4) 使用 MALDI-TOF/TOF MS 比較各 HA1 isoforms 之醣質差異,可發現等電點越低,也就是越酸之 HA1,其 m/z 1867.2 之醣鏈含量比例越高。 5) 未來將繼續以 MS 檢定 HA1 上面的 N-linked glycans。 |
| 09/02/25 |
4 林 之 儀 L-SP 的 PI 催化活性機制 |
|
|
|
| summary |
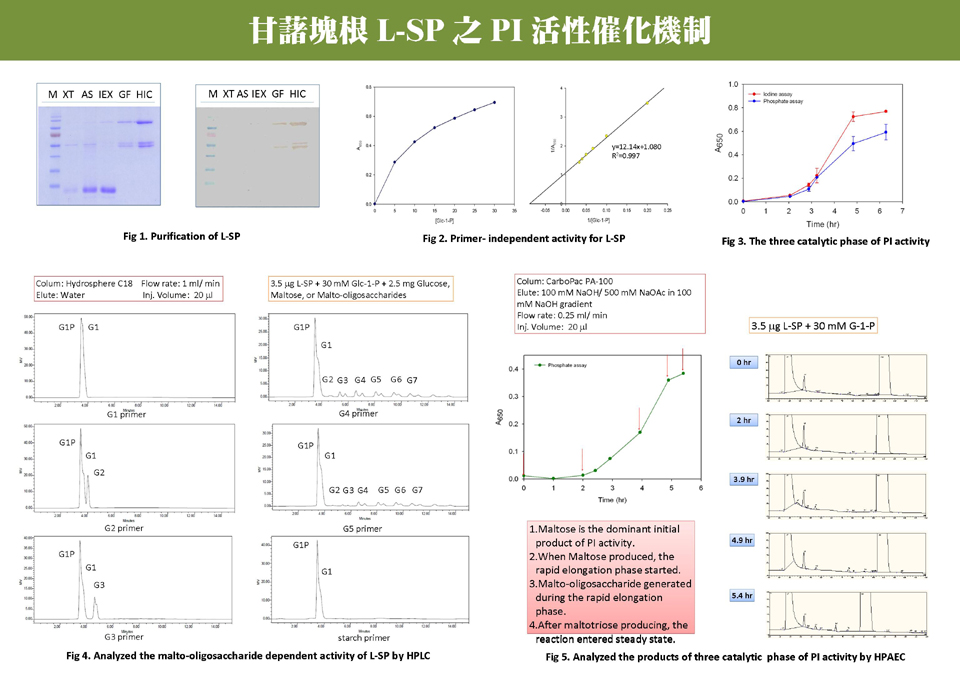
結果摘要︰ 1) 繼續進行 HPAEC,注意其結果需有再現性。 2) Glc-1-P 的 peak 可能擋住五醣及六醣,將使用 BaCl2 沉澱法嘗試去除。 3) 是否能將 injection volume 增大,把 HPAEC 的圖譜做漂亮一點。 4) 做長時間反應,測定活性的時間不能太長,否則會影響結果的可性度。 5) 每一張圖都應該要有 error bar。 6) 跑 IEX column 時要確定膠體已平衡完全,並且在一開始流洗時,就要收集 unbound 部份。 7) 反應產物可以用代謝體學專用的小分子 LC/MS/MS 分析。 未來工作︰ 1) 以 GC/MS 或 LC/MS/MS 來分析 L-SP 反應之產物。 2) 繼續進行 HPAEC,並加入 phosphate 為標準品,以得知反應後 phosphate 在圖譜之位置。 3) 使用 galactose 做為基質,看其合成是否與 phosphate 有關。 4) 進行 H-SP 之純化,並進行其 PI activity 分析。 |
| 09/02/11 |
5 眭 毓 庭 PCS 的活性機制探討 |
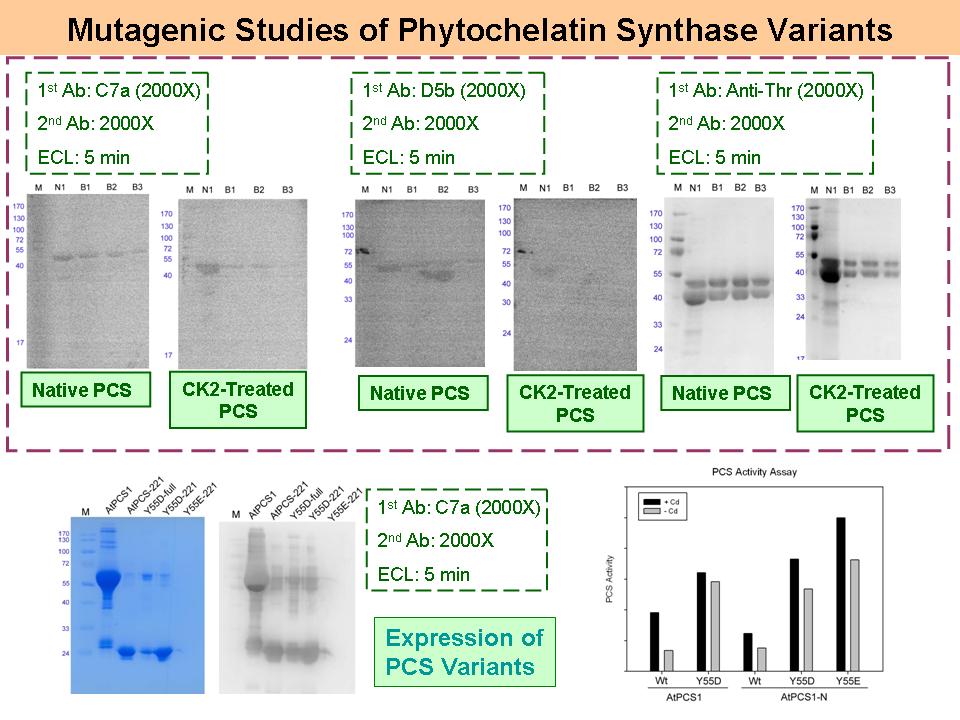 |
|
| summary |
1) PhosPRO phosphoprotein purification kit 純化到的是阿拉伯芥中所有磷酸化蛋白質,其中只有部分為磷酸化 PCS,需用 2D 電泳才能真正分析磷酸化前後抗體 C7a 的辨識狀況。 2) 必須留下純化前的樣本一起跑電泳及 Western blot,且蛋白質要定量跑電泳,比較其消長變化才有根據。 3) Activity assay 應該用相同的莫耳濃度來比較,而非以相同的蛋白質量。 4) Activity assay 中沒有加 Cd2+ 的對照組,應添加 EDTA。 5) 將所有現有的突變株,包括 Y55A、Y55E、Y55D 都一起來做活性分析。 6) 可以考慮再做 Y55H,預測活性應該會更強。 |
| 09/01/14 |
8 林 怡 岑 澱粉磷解脢 L-SP 與其他酵素的交互作用 |
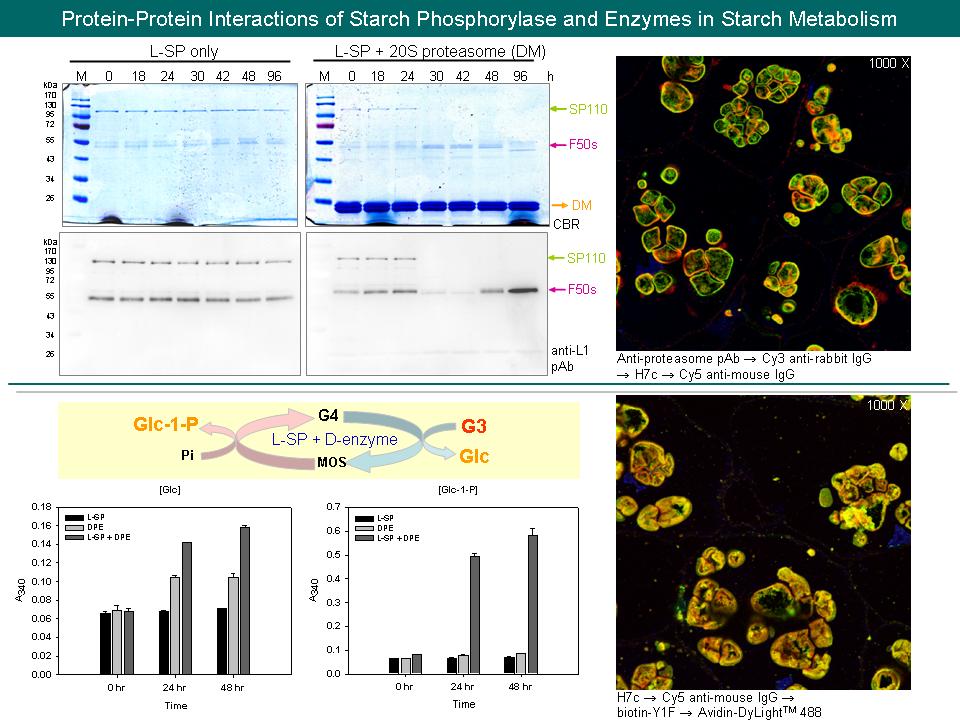 |
|
| summary |
1) In vitro degradation assay 可與張老師實驗室合作,設計實驗比較 20S proteasome wild type 與 deletion mutation 降解 L-SP 之差異性。 2) 利用 anti-ubiquitin Ab 偵測粗抽液中,L-SP 是否有 ubiquitination 的現象。 3) 購買連結螢光之 anti-IgM Ab 以偵測 J3b。 4) 比較直徑小於 5 mm 與大於 50 mm 之甘藷塊根中,L-SP 與 proteasome 之組織定位圖。 5) 利用 anti-L1 pAb 進行組織免疫螢光染色,觀察 L-SP 在不同時間之 45°C 加熱切片中,是否有階段性降解的現象。 6) HPLC 圖譜中,為何 maltotriose 以上之寡糖標準品,都會分裂為兩個 peaks? A) 可能是 a 與 b form。 7) 在給予 maltotriose 為基質的情況下,L-SP 與 D-enzyme 之酵素反應會產生 maltose 嗎? A) 可能有雜蛋白污染 (例如 b-amylase)。 8) Anti-20S proteasome 抗體是兔子傳統性血清,如何確認其是否具有專一性? A) 目前已用甘藷塊根粗抽液之全蛋白質來進行 Western,結果只偵測到一條 proteasome 色帶,無其他蛋白質色帶出現,故此抗體應具有專一性;另外也可進行免疫沉澱法,將所有沉澱之蛋白質進行 LC-MS/MS。 9) Proteasome 或 D-enzyme 在甘藷塊根中,蛋白質含量應該沒有 L-SP 多,為何螢光訊號那麼強? A) 由於抗體有單株、多株的差異,以及二抗或 biotin-avidin 放大訊號程度不同等問題,使得我們無法從螢光訊號強弱,來觀察蛋白質的實際含量差異。 10) Confocal microscopy 螢光訊號重疊之實驗結果能直接證明兩個蛋白質互相結合嗎? A) 由於放大倍率不足,螢光訊號重疊只能說明兩個蛋白質有 co-localization 的現象,還需配合其他蛋白質互相結合之實驗結果,才能推論兩個蛋白質互相結合。 11) 可進行 yeast two-hybrid 來佐證蛋白質之間的結合關係。 12) 一般 proteasome 大多位於細胞的哪一部分? A) 有 paper 顯示在細胞質、細胞核以及造粉體中都有發現。 13) 為何 HX 是 L-SP 加 proteasome,也是 L-SP 加 D-enzyme? A) 目前推測 HX 中可能含有不只一種形式之蛋白質複合體,而由原態電泳圖中,的確可觀察到多條 L-SP 高分子量活性色帶。 14) 在 L-SP 之 primer independent mechanism 中,D-enzyme 可能扮演的角色為何? A) 假若 L-SP 無法將形成的 primer 釋放出,D-enzyme 可能進行糖分解反應,幫助 primer 釋放,或者當 L-SP 延長 primer 到 maltotriose 時,交給 D-enzyme 進行後續的糖鏈延長作用。 |
| 09/00/00 |
1 報 告 者 主題 |
|
slide |
|
| summary |
|
{kind=link}