|
進度報告摘要 |
|
| 12/10/25 |
1 陳 又 楷 B2m 蛋白質的純化與 2-DE 檢定 |
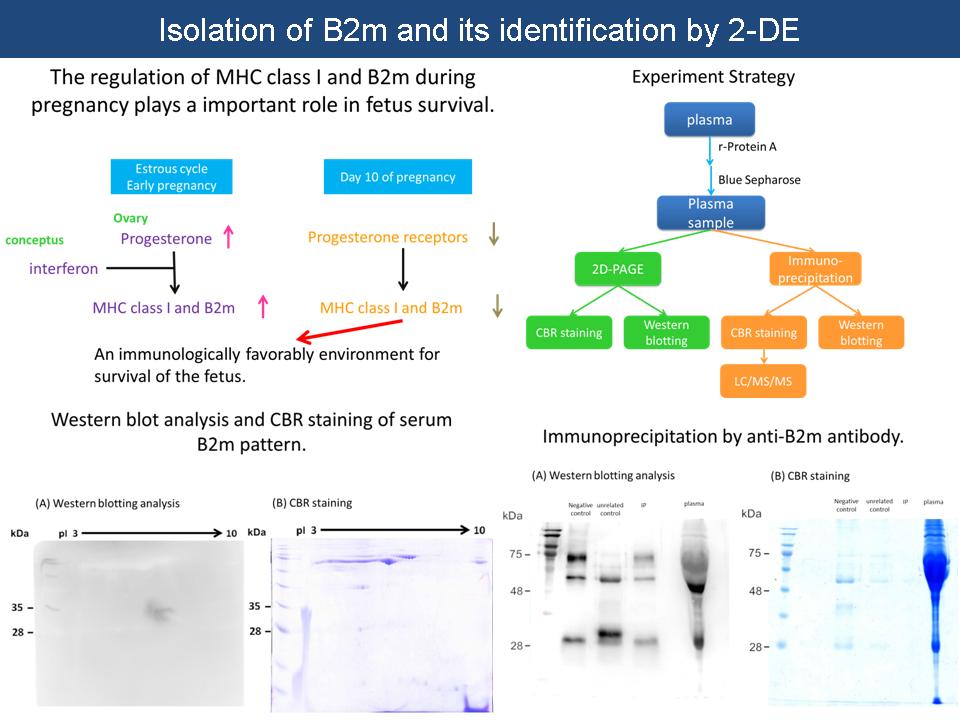 |
|
| summary |
(1) 為何二維分析結果無法呈現 B2m 之正確 pattern? 由於 Western blotting 結果尚未穩定,多次實驗皆未觀察到 B2m 之連續位點分布,需改善抗體濃度及反應時間,增加實驗操作熟練度。另外,直接電泳分析血漿樣本,觀察 CBR staining 和 Western blotting 結果並比較高產及低產豬差異。 (2) 免疫沉澱之原理及步驟如何? 利用抗體和抗原間專一性結合,自樣品中純化出目標蛋白質。是將抗體架接於磁珠,加入樣本並洗去雜蛋白,溶離後得到目標蛋白質。 (3) 目前免疫沉澱之結果? 由於抗體與磁珠架接效果不佳,溶離時抗體與 B2m 一起脫落,Western blotting 出現 3 個明顯訊號 (75 kDa, 50 kDa, 25 kDa),應為抗體及其斷片,遮蔽 B2m 位點。改善方法為:提高抗體濃度和延長架接時間,增加清洗次數以減少非專一性結合。 Future work: 完成生產預測實驗,並進行 B2m 定序,探討位點 1 和位點 2 的後修飾差異,以及其所代表的意義。 B2m 的分離一定要做好,並且有再現性。因此,親和層析的步驟必須仔細檢討,找出純化的最佳條件。 |
| 12/10/11 |
1 蔡 政 宏 Pho1-DPE1 複合體中澱粉磷解酶與 D-酵素之轉譯後修飾 |
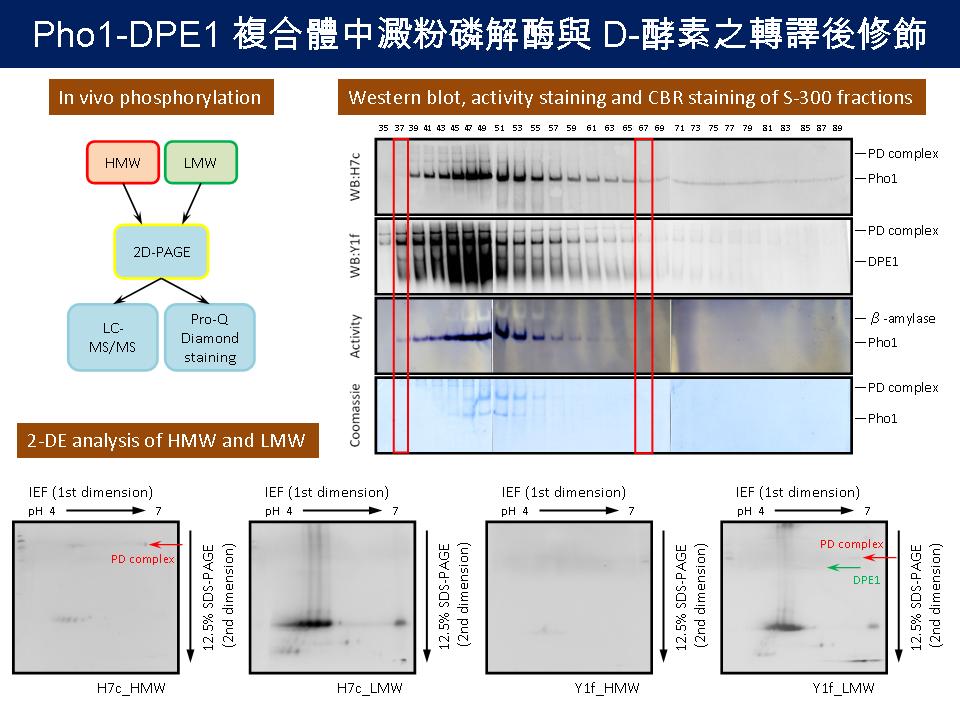 |
|
| summary |
進度摘要: (1) 將甘藷粗抽以膠體過濾分離之後,分別以 H7c 及 Y1f 抗體進行免疫呈色,決定 HMW 及 LMW 所在的分劃。 (2) 將 HMW 及 LMW 分別進行 2-DE,分析其圖譜差異。(未成功) 問答集: (1) 3 次都使用硫酸銨分劃? 硫酸銨分劃不穩定的原因? A 只有第二次採用。 不穩定的原因可能是 protein 樣本的量沒有固定,但可能還有其他未知的因素。 (2) 2-DE 未能成功的原因? A 在將蛋白質定量後再進行濃縮回溶的步驟,可能過程中會導致蛋白質流失。而且在選取 HMW 及 LMW 的樣本時,選取單一 fraction 本身蛋白質量就不多,不容易看到訊號。 (3) HMW 及 LMW 之定義? A HMW 為 SP 與 DPE1 形成的 complex,而 LMW 則是 SP 及 DPE1 在細胞中的 native form。 (4) 原本 2-DE 預期看到什麼樣的結果? A 預期應該會在同一個分子量看到依 pI 值分離之不同的點,證實有磷酸化的現象。 (5) 還要繼續使用硫酸銨沉澱法嗎? A 暫時先以濃縮管進行樣本濃縮的步驟。 (注意粗抽取液很容易把濃縮管阻塞!) (6) DPE1 和 SP 形成 complex 之機制? 實驗設計的理由? A DPE1 與 SP 可能是藉由磷酸化的方式形成 complex,因此實驗設計便是要證明 HMW 的蛋白質有磷酸化,LMW 則沒有磷酸化。接下來再證明將磷酸化的 HMW 利用去磷酸酶除去磷酸化之後,是否會恢復為 native form,便可證明磷酸化對形成 complex 是必須的。 (7) 如何直接證明蛋白質受到磷酸化? A 可以用 Pro-Q Diamond staining。 建 議: 活性染色解析力不足,可試採用梯度膠體電泳將 β-amylase 及 Pho1-DPE1 complex 分開,避免 β-amylase 干擾。 結 論: (1) 濃縮後先定量蛋白質,再進行膠體過濾,找尋最適合 HMW 及 LMW 分離的濃度。 (2) 先證實 Pho1 及 DPE1 會受到磷酸化。 (3) 要更加熟練實驗技術。 |
| 11/10/25 |
1 賴 欣 妤 追蹤 IFGBP3 以預測母豬之生產率 |
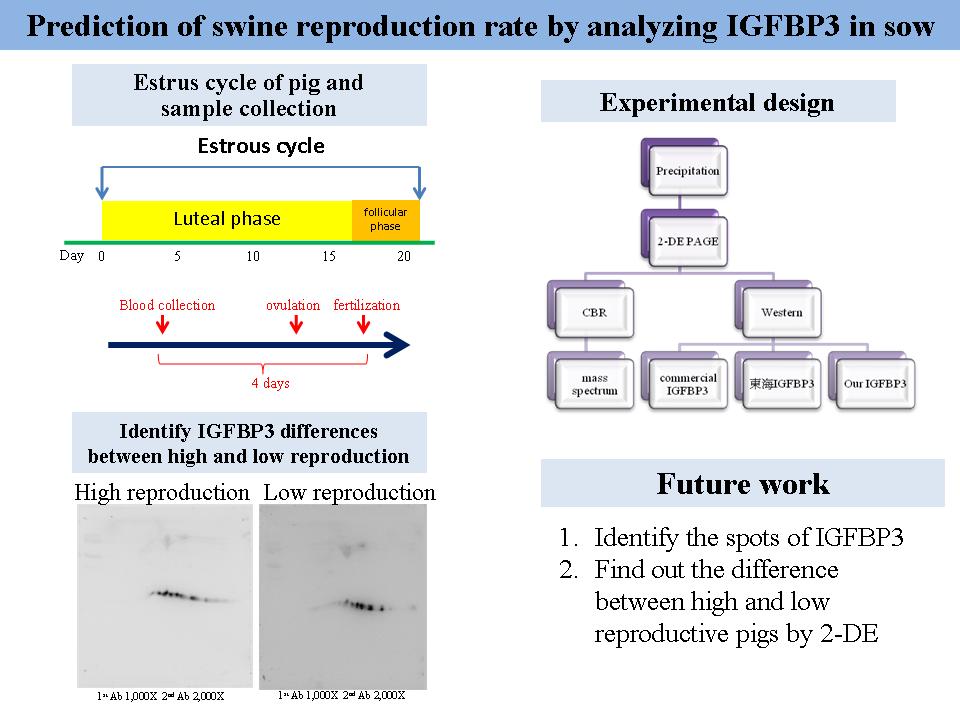 |
|
| summary |
1) 應介紹使用 IGFBP3 做為研究主題的緣起。 2) 調整 ppt 的順序,以增加報告的通順度。 3) 加入一張整理表格,包含 IGF-1, IGF-2, IGFBP3 的功能及特性。 4) 了解 IGFs, IGFBPs 的作用機制。 5) 要搭配參考 paper 的圖表較能說明 paper 內的觀點。 6) 是否要對 IGF-1, IGF-2 進行實驗。 7) 東海所做的抗體 (IGFBP3),是用哪一段 peptide 或是用 protein做 出來的? 8) 確認 IGFBP3 的抗體是否正確。 Comments: 除了對報告的方法與指正之外,應該多加強對研究內容的討論 (研究目標、方法、規劃等) |
| 11/10/18 |
1 胡 景 涵 搜尋癌幹細胞表面的標誌蛋白 |
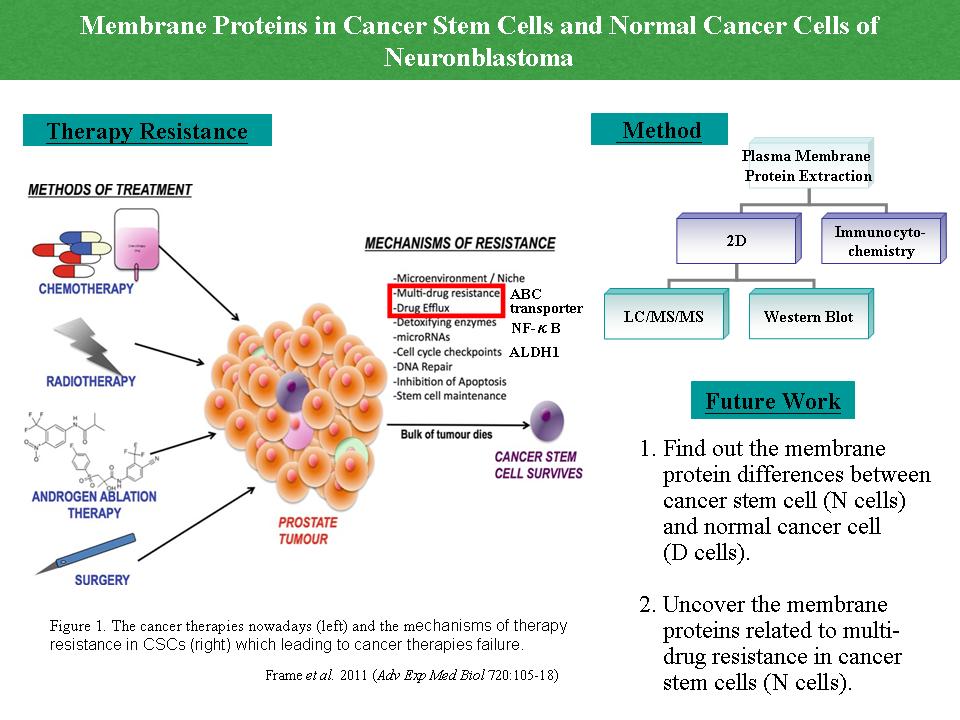 |
|
| summary |
1) 要說明清楚為何要做此實驗,實驗目標要說明清楚。 2) 每張圖要講清楚重點為何,尤其是實驗重點的圖,要將內容弄懂。 3) 找出 ABC-transporter 之保守性序列及其保守性。 (此 ABC 與下面的 ABC 不同) 4) 問清楚 ABC (avidin-biotin complex kit) 之價錢,可以用 ABC system 增加偵測靈敏度。 5) IEF, electrophoresis 直接以 2DE 表達。 6) 投影片之英文文法及報告時口頭禪要改善。 Comments: 除了對報告的方法與指正之外,應該多加強對研究內容的討論 (研究目標、方法、規劃等) |
| 11/10/11 |
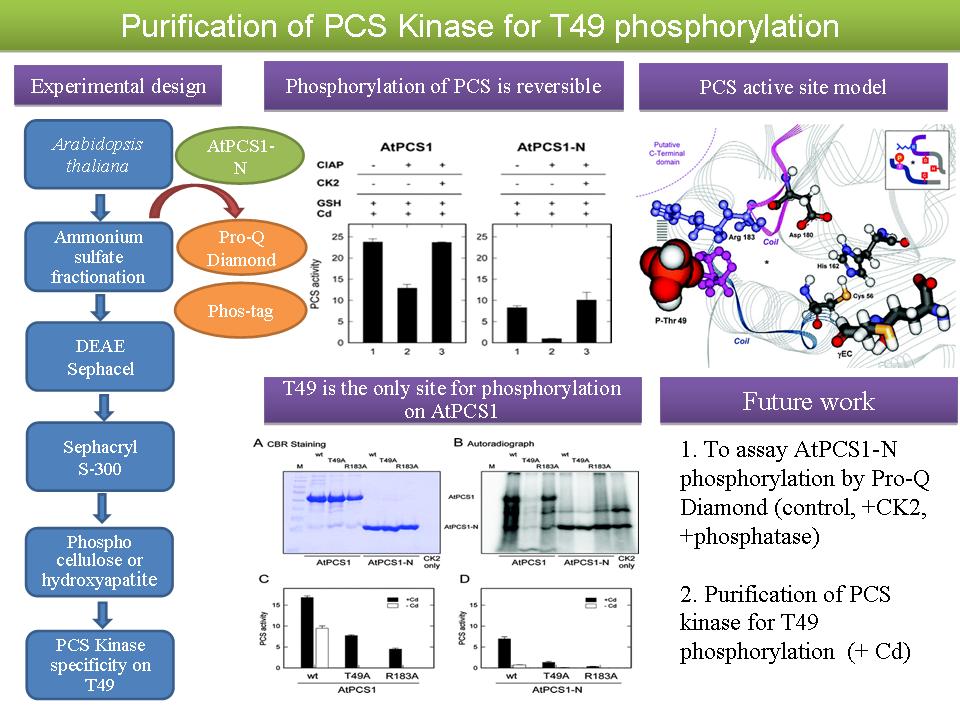
1 陳 怡 靜 PCS kinase purification and assay methods |
 |
|
| summary |
兩個取代放射線以觀察磷酸化蛋白質的方法: (1) Pro-Q Diamond stain kit 此方法可以將磷酸化的蛋白質偵測出來,一開始經過 Pro-Q Diamond stain 進行染色後來經過 excitation/emission visible-light scanning instrument 觀察是否有磷酸化蛋白質的存在,可使用於 SDS-PAGE 以及 2D gel。此產品和 SYPRO Ruby dye 並用,可以進行蛋白質的定量,更加了解蛋白質磷酸化的情形。 (2) Phos-tag 此新技術 2010 年由日本 Wako 公司研發,利用金屬離子 (Mn2+) 和 polyacrylamide 的鍵結,可以和磷酸化的蛋白質結合,不同的金屬濃度可以使磷酸化的蛋白質分離的更加明顯。此試劑只要在配製 acrylamide 溶液內加入 Phos-tag 即可,但此產品非常貴,每一次使用需要 1,773 元。 討論摘要: (1) 所用的 PPT 每個圖表都必須深入了解,並且有條理的向聽眾清楚說明。 (2) 要注意書與打字寫格式,英文的拼字、發音與用法一定要正確。 (3) 練習增加邏輯的能力及口說能力。 (4) 如何判定所純化的 kinase 就是 PCS kinase? (5) 磷酸 (phosphate) 不要說成『磷酸根』,因為『根』是 residue 的意思。 (6) 深入了解所有試劑的化學原理,以便順利執行實驗。 |
| 11/04/19 |
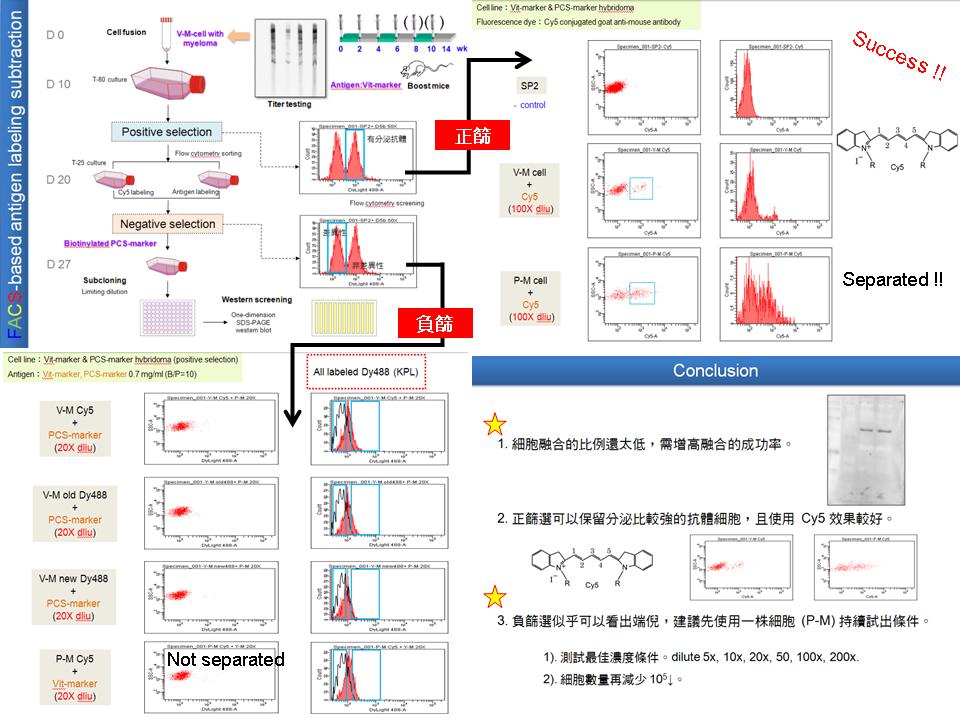
4 錢 思 翰 利用細胞表面分泌抗體的能力分離細胞群 |
 |
|
| summary |
1) 論文要將 model 1, 2 和整體 hypothesis 先說明清楚。 2) 正篩過程中抗原標定是否為同一時期 ? Ans: 是的。 3) Vit 是什麼 ? 若為 vitellogenin 則縮寫為 Vtg。 4) 目前先把負篩遇到的問題解決,斑馬魚樣本及小鼠才有機會做出來。 5)上一次流式取多少細胞 ? Ans: 約為 105 cells/tube。 6)下次應該要把全部的細胞都使用掉。 7) 目前遇到最大的問題是 fusion ratio 太低,除了隔天要換 medium 之外,離心力也很重要,融合後不能太大,也不能太小,就是 850 rpm (但要鏡檢)。再來就是融合瘤細胞 (SP2),在 T-80 養到 5, 6 成滿即可,避免細胞不健康。 8) 目前來說正篩選只需用 Cy5 就可以順利將細胞分群成有分泌和無分泌,但負篩選卻沒有辦法用抗原標定的方式分群,更深入的問題還有待調查。可考慮將抗原濃度做序列稀釋,針對細胞 binding 抗原的能力著手。 9) 可以將小鼠的採血做 2-DE 的 Western blot 測定效價。 |
| 11/04/12 |
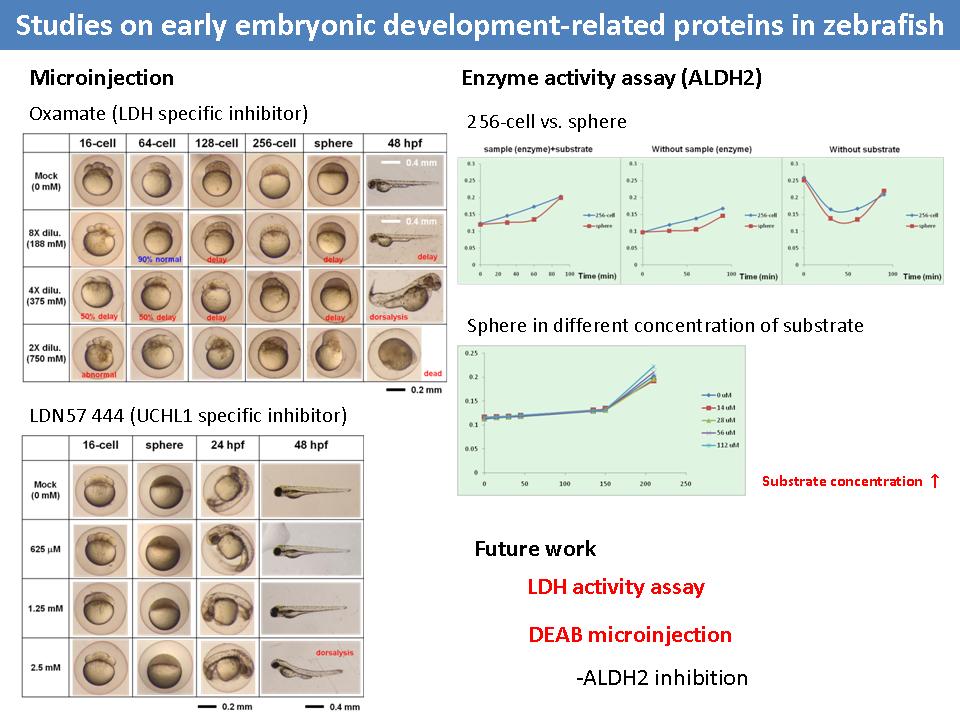
4 施 驊 珊 斑馬魚胚胎發育的相關標誌蛋白質 |
 |
|
| summary |
1) 以 oxamate 抑制 LDH 的結果發現,胚胎在 MZT 之前會發生生長延遲的現象;以 LDN57 444 抑制 UCHL1 的結果發現,胚胎要過了 MZT 之後才有不正常發育,且其 phenotype 不明顯。 2) Oxamate 的效果明顯,但應確定是否因為濃度太高所造成的影響,可以試打相同濃度且無毒性之物質。 3) 為了能證明各個抑制劑的效果,需以活性分析法測定 microinjection 後的胚胎酵素活性。 4) 查清楚是否有測定整個 glycolysis 效率的方法,以支持 MZT 前期 glycolysis 不活躍的假說。 |
| 11/03/22 |
2 卓 越 主題 |
|
slide |
|
| summary |
|
| 11/03/15 |
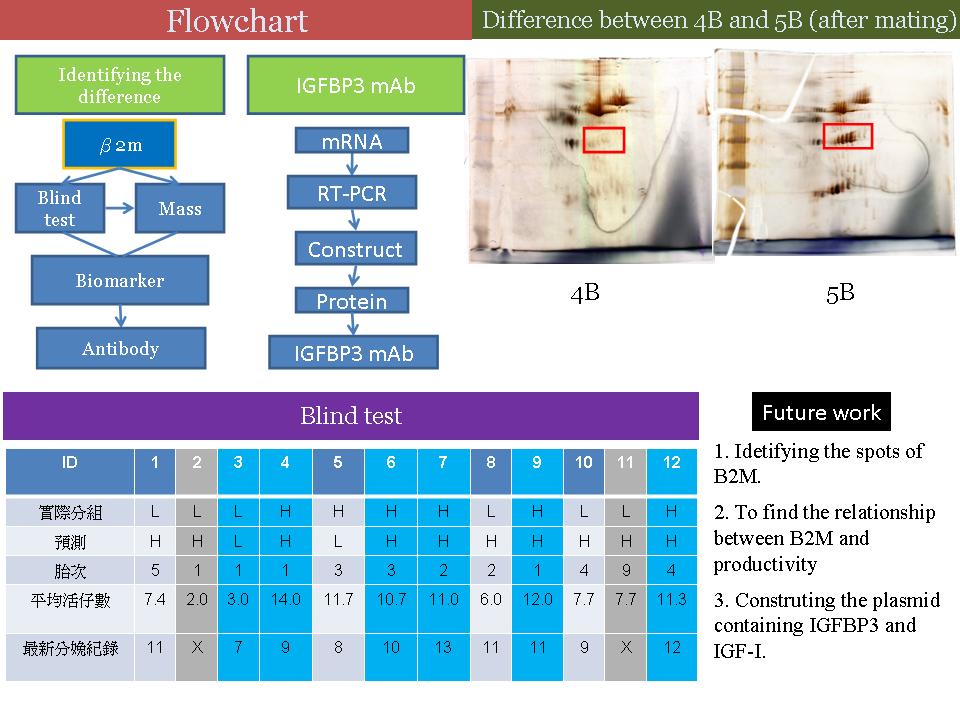
2 楊 己 任 找尋母源轉胚源過程的標誌蛋白質 |
 |
|
| summary |
目前成果: • 1) 目前以雙盲試驗來預測高低產結果還不準確 (38.8%),需找出 β2m pattern 與高低產的真正關聯。 2) 配種後的血清有明顯差異的部分,根據文獻指出極有可能是 IGFBP3 的位置。 3) 目前的實驗結果認為 IGFBP3 的抗體專一性可能不高。 建議: 1) 查 IGFBP3 glycosylation 情況,若有需要看能不能用實驗室的酵素切除,以便增加抗體專一性。 2) 不確定 IGFBP3 mAb 專一性如何,需再做 IGFBP3 Western blot,並借東海的 IGFBP3 抗血清以 Western blot 比對。 3) B2M 和 IGFBP3 在生理上的機制,如何跟得到的結果連結? 4) 讀過的 paper 可以開始整理摘要,做為論文的第一部份。 5) 在介紹時的故事,可以更完整連貫、有趣味性。 |
| 11/03/08 |
2 邵 子 瑜 AtPCS1 在植物中的表現 |
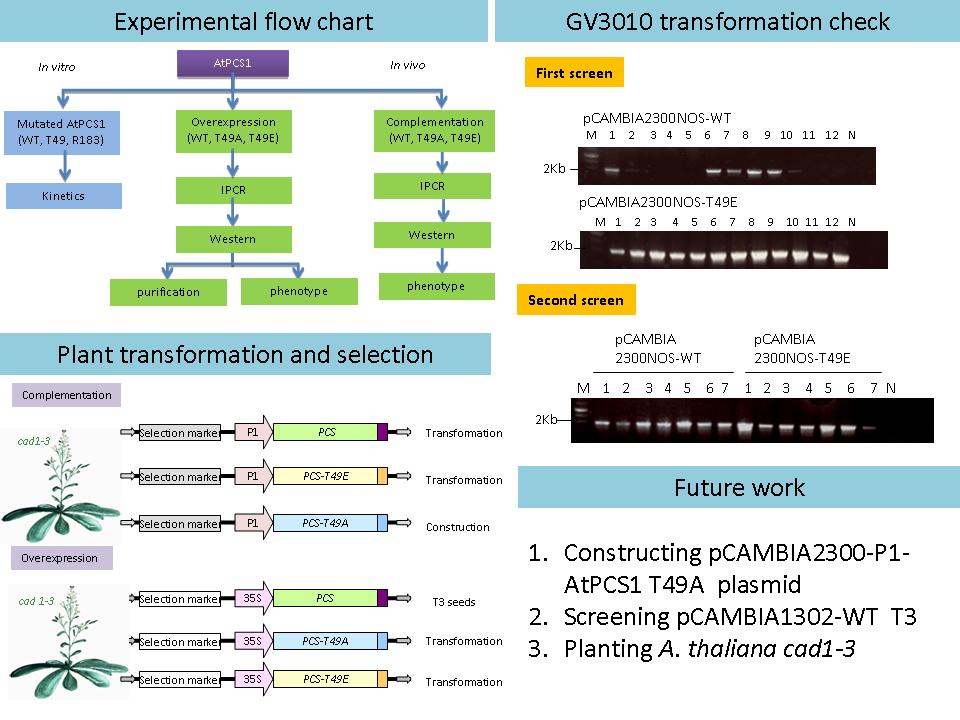 |
|
| summary |
目前成果: 1) 將 pCAMBIA2300-NOS-WT, T49E 以電穿孔法送入農桿菌勝任細胞 (GV3101),待進行阿拉伯芥 Cad 1-3 基因轉殖。 2) 對 pCAMBIA1302-WT, T49A, T49E 進行限制酶切割以確認片段大小,準備進行農桿菌電穿孔轉型。 建議: 1) 做到 T2 時可先做初步的分 析,以供 Master day 報告呈現。 2) 將 E. coli protein kinetics 改成 plant protein kinetics。 3) 製作未來進度之 schedule,以便控制實驗與時間。 |
| 11/01/25 |
1 謝 明 孝 禽流感病毒表面蛋白之表現 |
 |
|
| summary |
目前成果: 1) 完成 H6-HA1 五株單株抗體的腹水製備與純化。 2) Western blot 結果顯示 EB2-E5 單株抗體,無法偵測到 H6-HA1-P1 表現蛋白。 3) 使用 M1 表現蛋白測試了使用 peptide 免疫之 M1 單株抗體效價。 建議: 1) 第 44 頁要補上 DEAE 的管柱名稱。 2) 第 47 頁請畫出 HA-L 與 P2 基因序列的全圖。 3) 第三部份,目前抗體生產,腹水已相當足夠,但鼓勵多進行額外的生產研究。 4) 實際找出更重要的研究目標,先大量閱讀文獻並從中找出問題。 5) 將數據整理好才能看出實驗設計是否正確。 6) 查清楚哺乳動物細胞與重組病毒實驗申請的程序與規定。 7) 內容很多,但務必找出最終的研究主旨和焦點,以免失焦。 |
| 11/01/11 |
1 鄔 承 祐 CD133 (+) 細胞的癌化 markers |
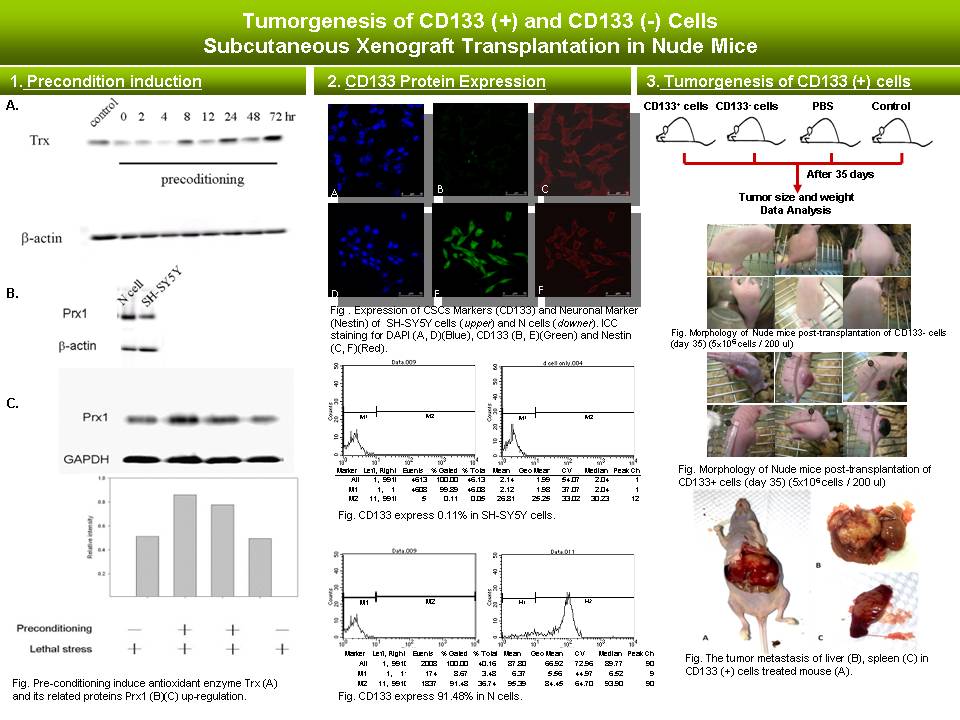 |
|
| summary |
目前成果: 1) Western blot 結果顯示 precondition induction 後之細胞株,抗氧化蛋白 thioredoxin 之表現有增加之趨勢。 2) Immunocytochemistry (ICC) 與 Flow cytometry (FCM) 結果顯示 precondition induction 後之細胞株表現 CD133 之癌幹細胞特有表面抗原。 3) 免疫缺失小鼠經皮下注射 CD133 (-) 與 CD133 (+),結果發現 CD133 (+) 之細胞株具有腫瘤生成與轉移之特性。 建議: 1) 何謂 ATP-Binding Cassette (ABC) Transporter ? 2) 簡介 Cancer stem cell marker CD133 相關資料。 3) SH-SY5Y cell line 來源與緣由? 4) Nude mice 的免疫缺陷為何? 5) 何謂 side population? 6) Thioredoxin 與 Glutathione 是否有其相關性? 7) 簡介 Hoechst 33342 DNA 染色法。 |
| 10/12/14 |
12 林 怡 岑 L-SP 與 20S proteasome 的交互作用與與功能 |
 |
|
| summary |
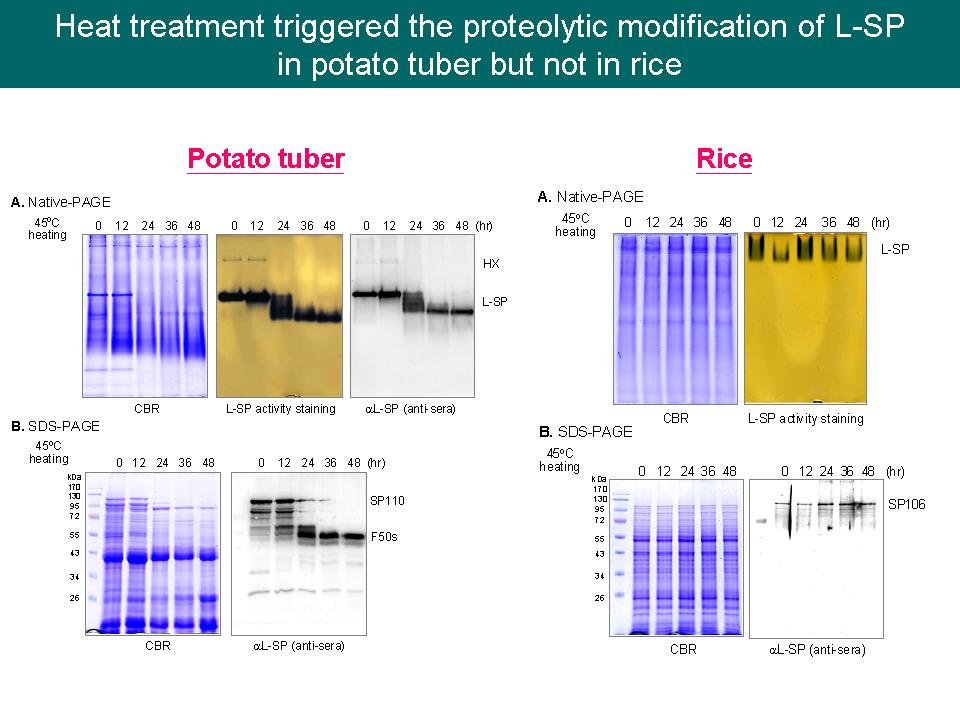
實驗結果: 1) L-SP 與 20S proteasome 之結合顯示 L-SP 可能經由一個不需 19S、ATP 與 ubiquitin 之 20S proteasome 路徑降解 L78,而且此降解作用會因 45C 熱處理而加速進行,此降解性修飾可能改變 L-SP 的生理活性,由合成澱粉轉為磷解澱粉,產生 Glc-1-P,進而促進 NADPH 生成,來減少因熱逆境產生 ROS 所造成的細胞傷害;另外,此降解現象也在馬鈴薯塊莖中觀察到。 建議事項: 1) 如何給予甘藷切片 MG132 處理?有無滲透到組織中?處理完有無沖洗? Ans:甘藷塊根切片 (2 mm) 於 45C 熱處理前,先浸泡在 100 mM MG132 下室溫反應 2 h,整個實驗流程是參考 Fettke et al. (2010) New Phytologist 185: 663-675。 2) Satoh et al. (2008) 與 Hwang et al. (2010) 是用稻米的哪一部份抽取 L-SP,有無分胚乳或胚芽? Ans:他們所使用的分別是用胚乳 (去除胚芽與種皮) 純化之 L-SP 以及 E. coli 表現之 L-SP。 3) 我們所用的稻米之來源與品種? Ans:由 518 實驗室所提供的台農 67 號,成熟曬乾的種子。 4) 由演化樹是否能觀察出甘藷、馬鈴薯之 L-SP 與稻米中的 L-SP 有何不同? Ans:它們分屬於雙子葉植物與單子葉植物。 5) 可探討單子葉與雙子葉植物的醣類代謝差異,找出 L-SP 之降解性修飾為何在這兩種植物中會有差異。 6) 一般給予熱逆境後的生理反應是什麼? Zeeman et al. (2004) 他們是如何給予阿拉伯芥逆境的? Ans:Heat shock proteins 表現量上升、水分含量下降、膜系不穩定、以及 ROS 生成。 Zeeman et al. (2004) 所給予的逆境是改變植物的生長濕度。 7) 在開始寫 Discussion 前要多看 paper 比較,不錯過任何一篇重要 paper,寫完一個觀點就可以先給老師看。 8) D-enzyme 有測定活性的方法嗎?可比較 monomer 與 complex 形式的活性差異。 Ans:可測定 Glc 之釋出。 9) 可用抗體偵測 D-enzyme 和 L-SP 之間的 binding site,抗體抓不到的位置可能就是互相結合的位置。 10) 若要討論 SP-DPE complex 所組成之結構,必需重複瑩貞的實驗結果,使結果更清楚。 11) 若 SP-DPE complex 結構中 D-enzyme 位於內側,那代謝機制是否為先由 L-SP 作用,再將產物交給 D-enzyme? 12) FRET 結果要有 negative control,可用較成熟的甘藷塊根切片進行實驗。 13) 若要觀察 SP-DPE complex 所形成的高分子量色帶,光看 native-PAGE 可能不足以證明 (色帶太雜)? Ans:會挑選幾個生長時期,進一步利用 gel-filtration 來觀察。 14) 需釐清 SP-DPE complex 之代謝作用機制,以及此 complex 形成的目的為何。 |
| 10/11/30 |
14 何 杰 龍 HA1 上糖類部份對免疫抗原性的影響 |
 |
|
| summary |
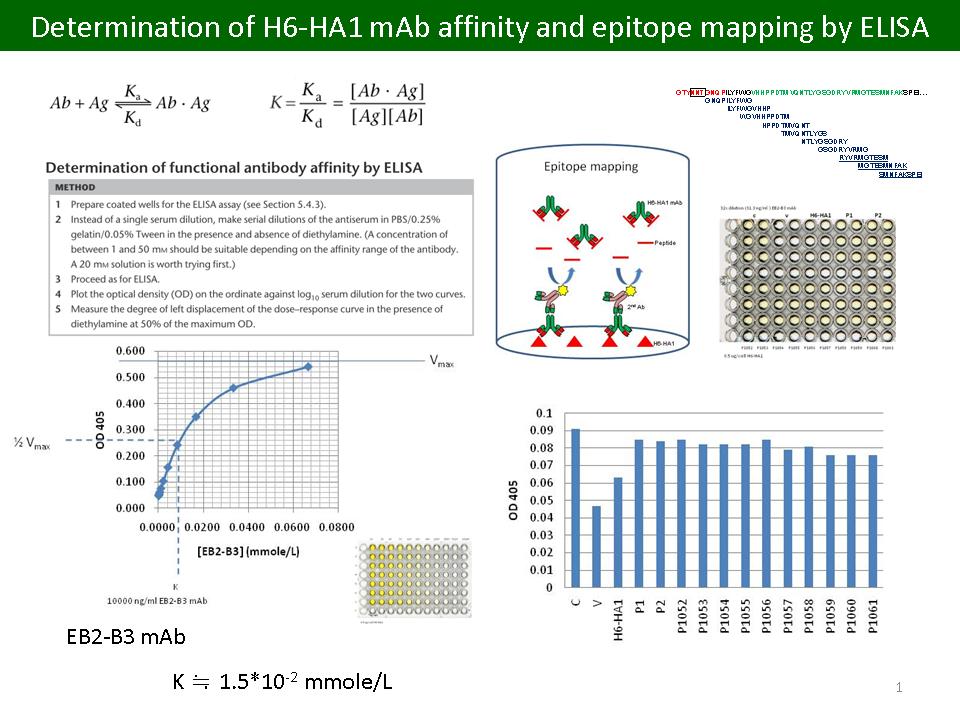
1) 本實驗室 H6-HA1 單株抗體具特殊性質 - 當醣蛋白質 HA 上的醣類被去除,此抗體與 HA1 結合的能力會減弱,甚至消失。這是整個研究架構的開端,據此與後續實驗提出『HA 結構保護機制』的假說。但 D Day 有老師建議必須釐清 deglycosylation 的過程是否會因 PNGase F 含某種蛋白脢而產生誤判?為此補 control:以 HA1 表現蛋白質進行deglycosylation,結果證實,我們所看到的現象的確由外加的 trypsin 所造成,並非 PNGase F 所含某種蛋白脢而產生的誤判。 2) 本單株抗體對 H6 亞型禽流感病毒 HA1 具有很強專一性,不過一直沒有量化。我們以 ELISA 進行抗體與抗原間親和力的評估,初步得到 K 值為 1.5 x 10-2 mmole/L。並以此實驗選定 312.5 ng/mL 的抗體濃度進行 epitope mapping。 3) 取 HA1 上可能為此抗體 epitope 的區域,設計部分重疊的 10 個片段進行 peptides 合成,以 bELISA 進行 epitope mapping。以 2838V 純化病毒 (V) 與表現蛋白質 (H6-HA1) 為 positive control,訊號皆明顯下降。可惜的是此批設計的片段 bELISA 結果都無明顯差異,已經設計第二批 peptides,目前正由廠商合成中。 4) 實驗與資料整理與撰寫論文的速度要加快,請列出論文發表流程的時間表。 |
| 10/11/23 |
3 林歆佑 PCS 在植物中的表現 |
 |
|
| summary |
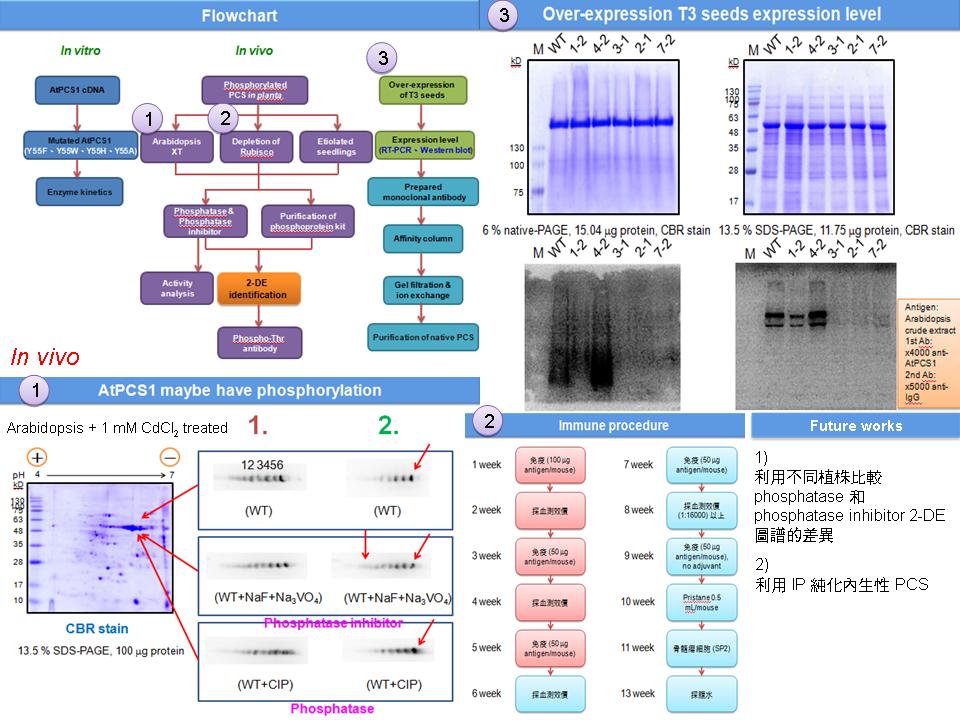
1) 重新測量 over-expression T3 轉殖株的 mRNA、protein 表現量和 phenotype 性狀的比較。 2) 測量 over-expression T3 轉殖株的活性。 3) 要放上次 Master Day 老師們所提的問題並檢討,以及前一次進度報告的摘要, 並且看學位論文後面的答問錄。 4) 要做 CIP 處理不同時間來比較 2D 圖譜的差異。 5) 要做實驗計畫把時間和目標訂出來。 |
| 10/11/16 |
3 錢 思 翰 差異性抗體庫的一些問題 |
 |
|
| summary |
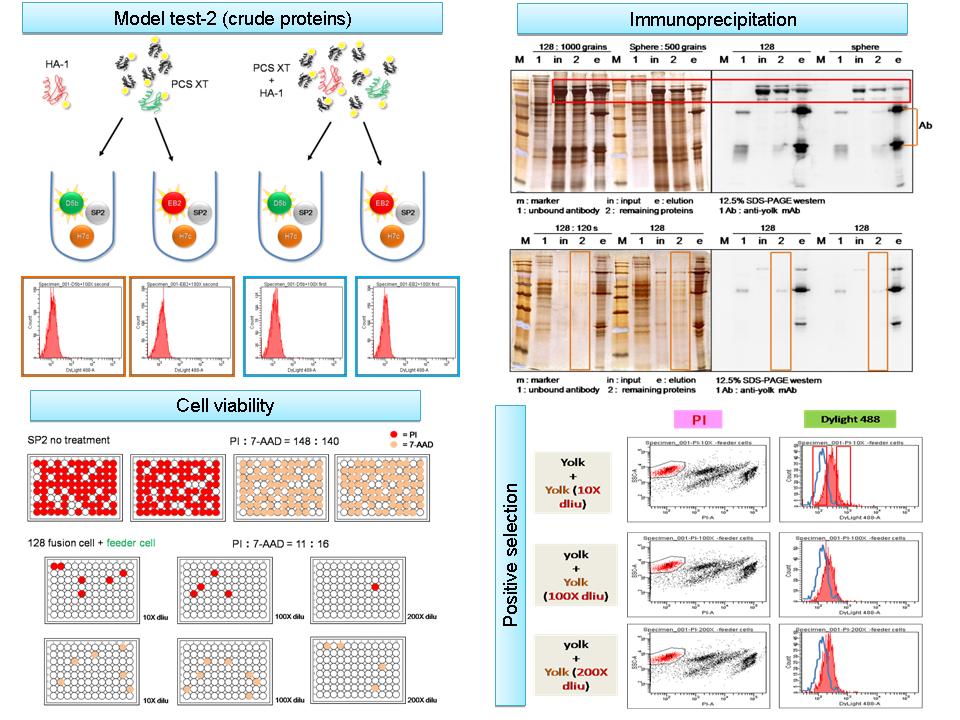
1) 做篩選的時候訊號 shift 的意思? 原本在 model 測試都很漂亮,可以分得出兩群很明顯的群落,原因是我們篩到的細胞都是分泌很強的抗體,表面都有嵌著許多抗體;事實上,取出小鼠的 B cell 是分泌有強有弱抗體的細胞,這樣平均起來,整體訊號還是很弱,當然沒辦法分出 peak,所以我們在負篩的時候才都篩到 yolk 的抗體,因為那只是效價較弱分泌 yolk 的細胞罷了。 2) 細胞分泌的抗體是否為 membrane bound? 因為 model test 有膜上抗體不代表 fusion 完後也會有。 我做了 yolk 的正篩以後,確定整體的細胞有螢光強度,跟 control 比有明顯的 shift,代表膜上是有的。 3) Briclone 效果? Fusion 後幾天做? 要細胞成熟後分泌抗體在膜上 14 天是否太早? 也許可以分兩盤 T-80。 Briclone 強調是可以增加分泌抗體的能力,細胞融合完後介於 10~16 天最好,生長曲線最好也還沒長滿競爭。目前的方式就是以分兩盤做。 4) 染 7-AAD 時不能看 PI 的訊號,會有互相干擾,不準確。 5) PI 和 7-AAD 比較完後選擇 PI,但是分選過程中還是可以把 PI 的量減半,看起來 PI 的訊號還是很強。 6) Model test-2 目前只做一次,實驗再現性不夠,須重複。如果還做不出來可以用 (純蛋白 biotinylated) 混入 (PCS crude extract 未標定),看是不是粗抽取物中的干擾問題。 7) 下次再做 model test-2 要設計好實驗,control (+) 要一起做。 8) 集合所有的 beads 成管柱,將反應 sample 的比例放大,一次處理較多的樣本,因為 beads regeneration 的能力較差,用幾次就沒什麼效果。 9) Immunoprecipitation 完後的蛋白質濃度高低? Sample lost % 若干? 在同一管反應體積沒有放大或縮小,yolk 有被明顯的吸附,可能會使整體蛋白質濃度下降,不過成功的有吸附掉 50% 以上,剩餘的蛋白質 band 還是可以看得到,也許會 lost 掉一些蛋白質,但這是不可避免。 10) 吸附完後的 sample 會有殘餘的 IgG,是小鼠血清內的,在免疫應該不會有太大的影響。 |
| 10/11/09 |
3 施 驊 珊 搜尋胚胎發育期間的蛋白質 markers |
 |
|
| summary |
1) 如何知道在小鼠的 oocyte 中不需要 glucose phosphate isomerase ? 在 2002 年 Kelly等人研究發現 glucose phosphate isomerase 基因被 knockdown 後,依舊可以生長到,代表此時胚胎的代謝來源並非 glucose。 2) ALDH 與 MMSDH 對 branch-chain amino acid 結構是否有專一性 ? 此外,須了解 ALDH 與 MMSDH 在其中的機制。 3) 若要製備抗體,是否製作豬的蛋白質,可利用在比對高產低產差異。 4) 目前先以半定量實驗為主,確認 mRNA 表現量是否跟 protein 結果一致。 |
| 10/10/26 |
1 卓 越 L-SP 的選殖與表現 |
 |
|
| summary |
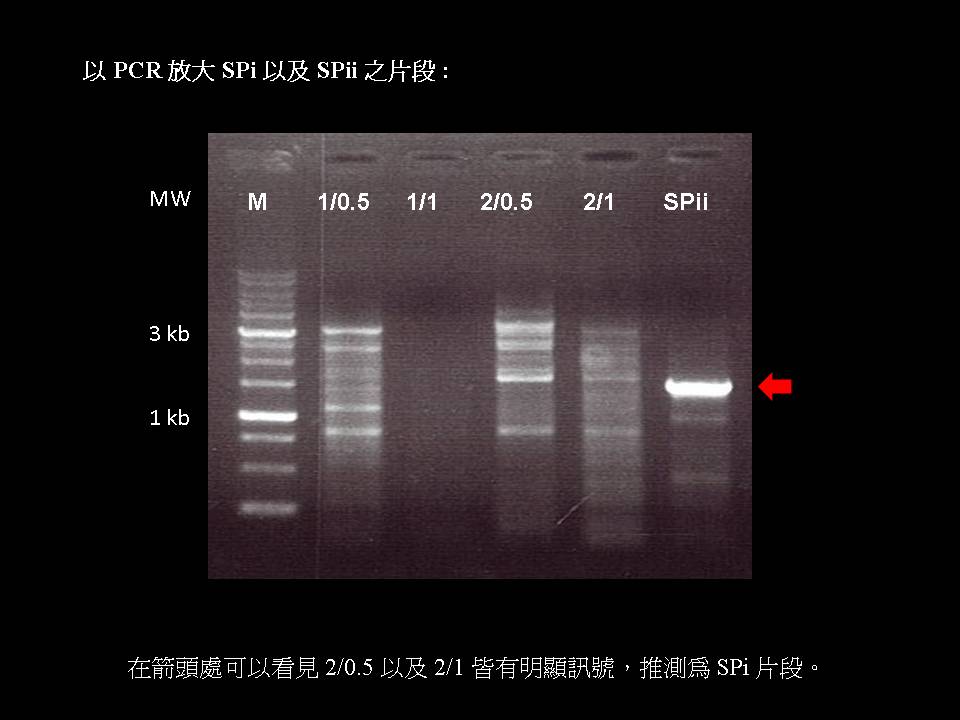
1) SP 序列中的 L78 位於 cDNA 上 1,239~1,320 鹼基對的位置,即整段 L78 皆位於 SPi 的 3’ 端。 2) 經過測試 PCR 程式的各個溫度之後,決定選用 57ºC 在 cDNA/primer 為 2/0.5 的條件進行 PCR,以取得較高純度,但較低濃度的 SPi 作為下一 PCR 循環的模板股。 (因為使用 57ºC 時較 56ºC 有較單純卻較淡的條帶) 3) 未來將以含有限制酶切位的引子進行第二階段的 PCR,再放大 SP 序列的量之後,進行 TA cloning。 4) 除了改變 cDNA/primer比例及溫度外,嘗試不同的 PCR 程式,例如提供兩個引子沾黏循環,以加強引子的附著。 5) 設計不同的選殖策略,例如將不好選殖的 L78 從中間分成前後兩段選殖。 |
| 10/10/19 |
1 楊 己 任 尋找母豬胚胎 MZT 轉換的生物標誌 |
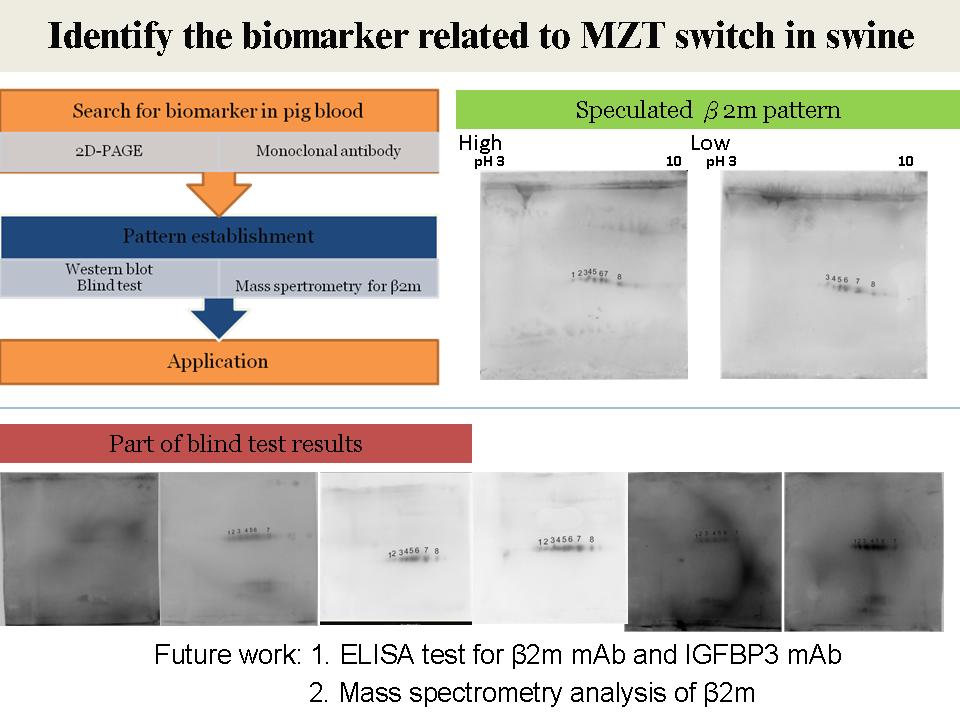 |
|
| summary |
1) 要找一個合適可信的 internal control (i.e. tubulin) 及定量方法。 2) 整體口頭報告,切入主題時不太連貫。引用文獻,需寫出期刊名。 3) 找出 β2m 的序列重做抗體。 4) 可用免疫沈澱找與 β2m interaction 的蛋白質。 5) 針對 β2m 的生物資訊學探索 (data mining) 加上 paper research 搜索 β2m 所知的相關生理功能。 |
| 10/10/12 |
1 邵 子 瑜 植物螯合素合成酶 PCS 之 T49 突變株磷酸化分析 |
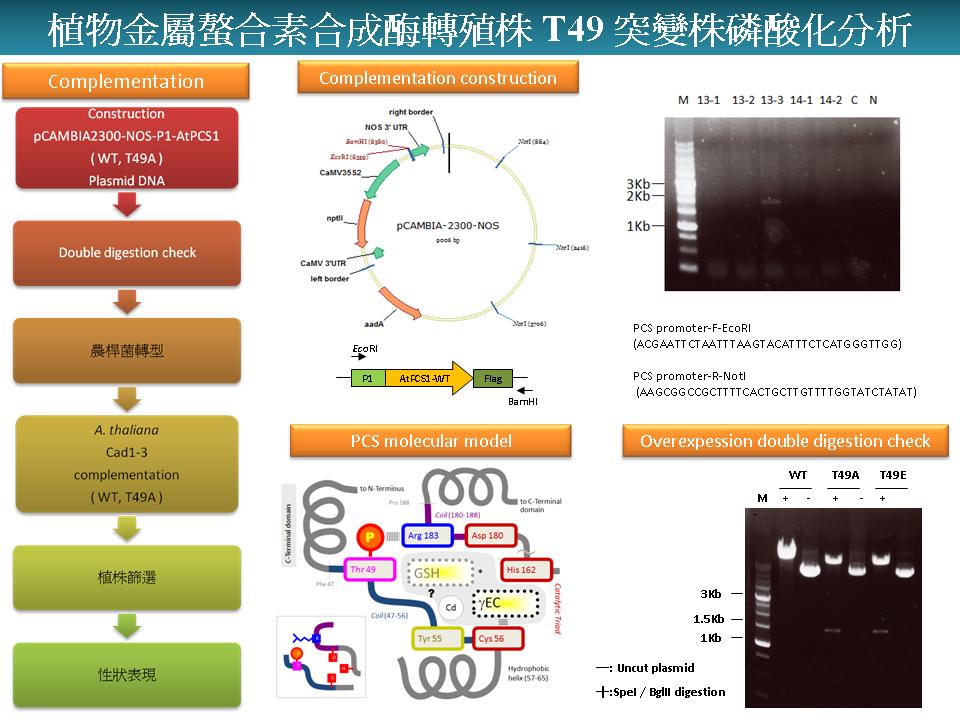 |
|
| summary |
1) 未來工作要集中在 complementation 實驗,以證實 PCS 的抗重金屬功能。 2) 要注意一般書寫格式,以及投影片上正確使用英文文法。中英文要統一。 3) 投影片中的圖太小,請儘量放大。 4) 要清楚說明實驗目的與預設結果,整個論文才能有足夠的說服力。 |
| 10/05/25 |
5 謝 瑩 貞 L-SP 與 D-enzyme 交互作用與可能的生理功能 |
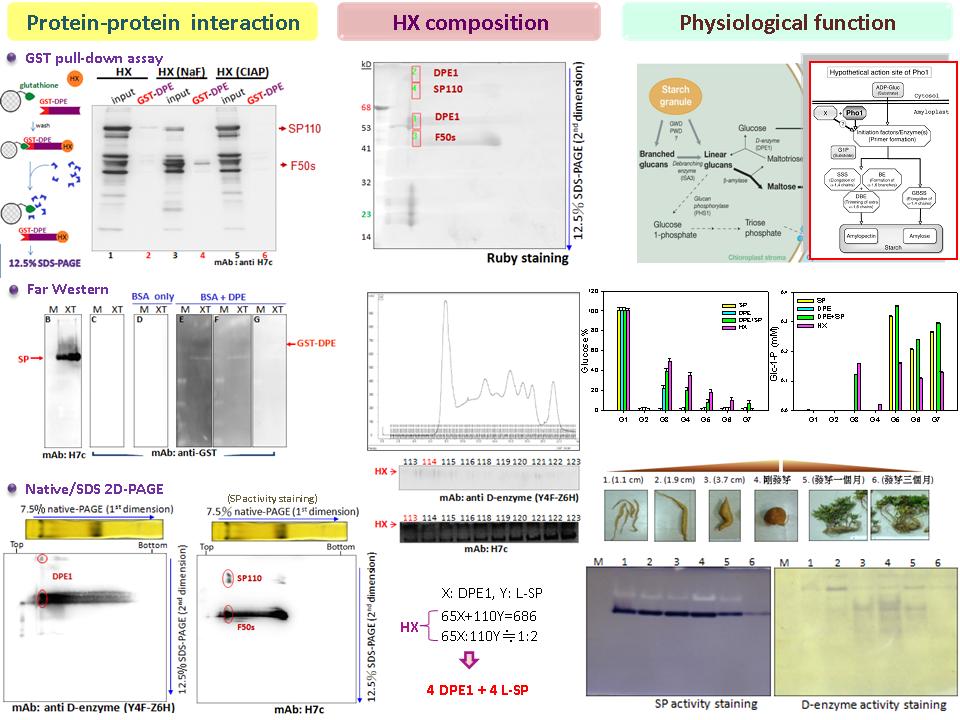 |
|
| summary |
1) G1P 一定要以 Glc-1-P 表示,口述時 G2, G3, G4, G5 應以 maltose, maltotriose, maltotetraose 以及 maltopentaose 說明,避免聽者混淆。 2) 未涉及 Km、Kcat,不能說是酵素動力學。 3) 設計實驗的目的及過程都需在報告時說明清楚。 4) 分析形成 HX 對 D-enzyme 或 L-SP 有何幫助。 5) 試著由結果推測出酵素作用的途徑,並進一步驗證反應過程中,是否有預期的中間產物生成。若欲證實 D-enzyme 可將兩個 maltotriose 轉變成 maltopentaose 給 L-SP 當受質,則可觀察反應過程中是否有 maltopentaose 產生。 6) 實驗都是設計分解的方向,應將實驗結果與推測的路徑作連結,說明 L-SP 如何參與澱粉合成。 7) 提升實驗結果的真實性有三個方法: a) 重複實驗,至少三次以上。 b) 做好控制組,因為控制組也可以說好故事。 c) 以其他證據支持實驗結果。 8) 找其他可以支持 HX 中 D-enzyme 及 L-SP 之次單元體比例為 1:1 的證據。 9) 整理實驗結果的工作固然重要,但最後必須將實驗結果分類,並強調實驗的重點。分析實驗結果後,應先整理出各小節的結論,最後再依照邏輯連結成完整結論。 |
| 10/05/04 |
5 黃 迺 茵 在阿拉伯芥過度表現 PCS 對植物鎘耐受性的影響 |
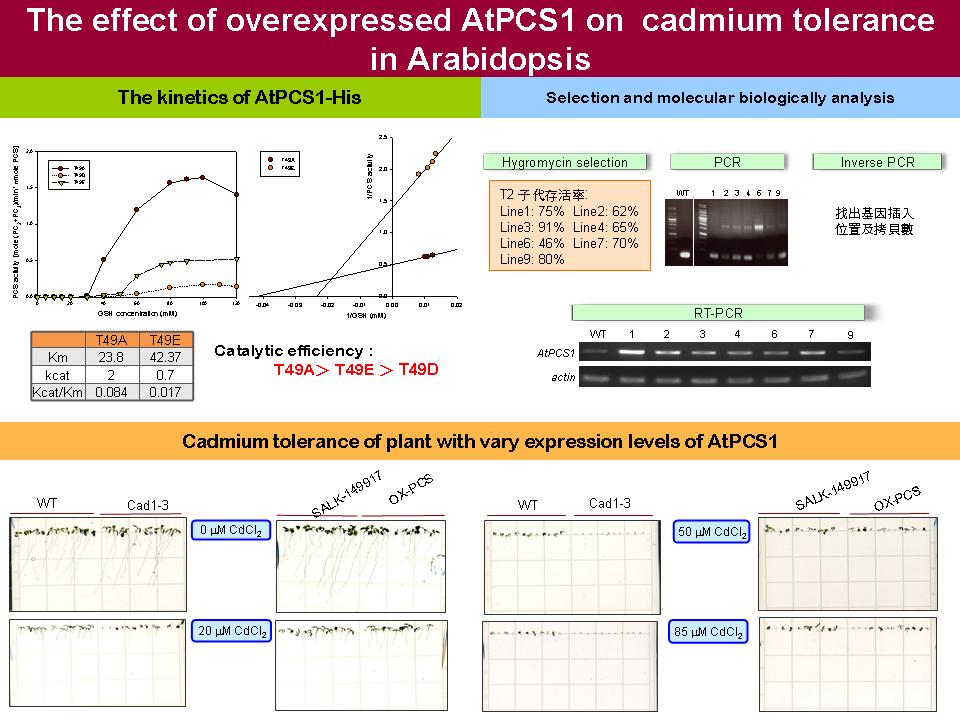 |
|
| summary |
1) 過度表現 PCS 轉殖株的 T2 子代已經過抗生素篩選,接著將以 IPCR 確定其基因插入位,再進行 PCR 確定此 T2 為 homozygote ,才能收取其 T3 子代,進行接下來的 PCS 性狀分析。 2) 每次實驗都要做好控制組,這次的酵素動力學未做 wt 當控制組,之後做三重複時將補做。此外,在 PCS 活性剛升起來的基質濃度應密集配點,才能看出是否為 sigmoidal 曲線。 3) 將 PCS 以 CIAP 去磷酸化後,其活性沒有明顯差異,但以 CK2 磷酸化後的活性確實有明顯提升。經 CIAP 處理的 PCS 活性之所以沒有明顯改變,可能是由於純化出來 PCS 重組蛋白本來就沒有磷酸化,或是已被水解掉。 4) 在 cad1-3 過量表現 PCS 的轉殖株經各種濃度鎘處理,發現可稍微降低 cad1-3 的鎘敏感性狀,之後將進一步觀察鎘耐受性是否跟 PCS 表現量成正比。 |
| 10/04/20 |
2 林 歆 祐 PCS 的活性區 T55 探討 |
 |
|
| summary |
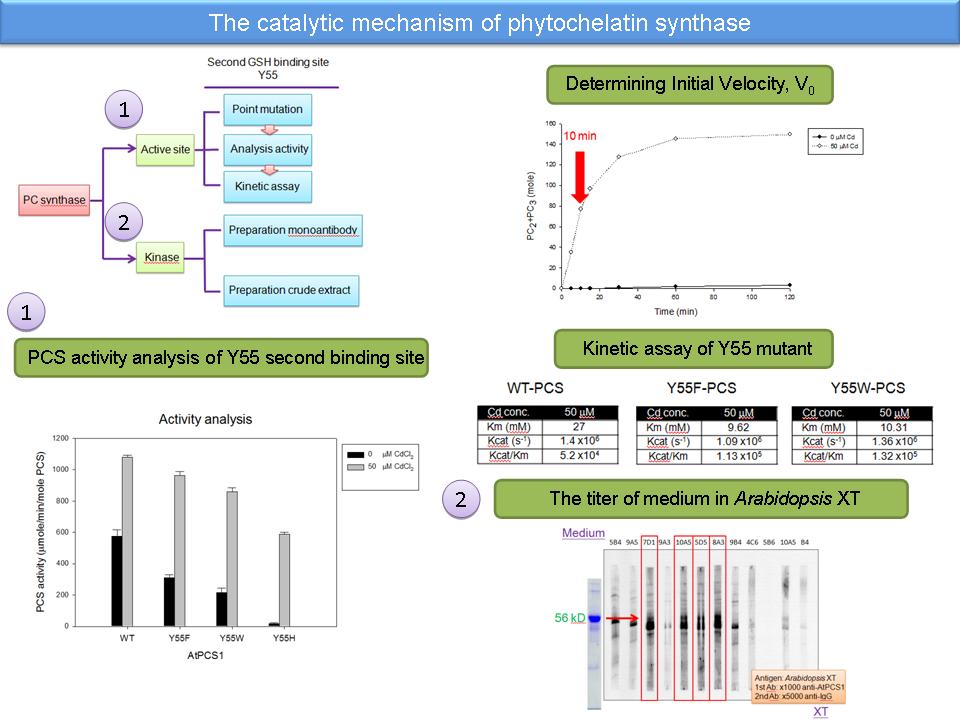
1) 以新條件做 Y55 mutant 的活性分析,並且要加做 Y55A 來比較 (也要做 wild type)。 2) 要再 repeat kinetic assay,確定其數據之可靠性。 3) 所用酵素定量要準確,每次都要以電泳色帶定量。 4) 從 kinetic assay 的圖可以看出也許有 substrate inhibition 現象。 5) 雙倒數的圖和酵素動力學的 Km 及 Vmax 測出來的結果不太一致,將換另一種方法去求出來比較。 6) 可能 Tyr 換成相同芳香族的 Phe 或 Trp 都可以恢復其活性。 7) Y55H 活性較低可能是因為帶有些微正電,而影響到鎘攜帶受質進入催化區。 8) 純化出來的單株抗體需要再做一次 limit dilution 以確保其專一性與純度。 9) Arabidopsis 的粗抽取液需要做簡單的純化步驟,以去除 rubisco 干擾。 |
| 10/03/31 |
2 錢 思 翰 斑馬魚胚胎蛋白質的抽取 |
|
|
|
| summary |
1) 不可能做出全部蛋白質體的抗體庫,因為蛋白質太龐大,只需比較差異的關鍵蛋白質即可。 2) mir-430 抑制何種基因的表現? 3) Paper 上的 1-D gel 用何種染料? 為何 9 和 26 hpf 的 patterns 差那麼多? 4) 可以使用簡單的純化把 yolk 去除掉 (gel filtration)? 5) 把每個純化的 fraction 跑電泳,自己選擇看是要 disc 或 SDS。 6) 純化出來的 yolk proteins,可以做看看抗體收集起來成 subtraction column。 7) 若是在免疫過程中 yolk 所產生出大量、非關鍵性的抗體該如何解決?差別性蛋白質所產生的抗原性較弱該怎麼辦? 8) 可以試試看 256 或 512 cell 也許 yolk 並沒那麼多,比較好分離,再和 sphere 做比較。 10/05/14 討論: 1) 一定要先去掉大量的 yolk 蛋白質,否則抗體庫很難達成。 2) 儘量做出抗這些 yolk 蛋白質的抗體,然後以免疫吸附法去除之。 3) 因為胚胎前期的蛋白質量很少,是否可以找比較後面而有意義的 stages 去比較? 4) 若魚卵不方便製備,是否可以借生科院的魚房自己培養? 5) 真的都無法成功,就得退回找 10 種模式蛋白質混合,然後比較 10 + A 與 10 + B 的測試。 |
| 10/03/24 |
2 施 驊 珊 斑馬魚胚胎發育期間的蛋白質體比較 |
 |
|
| summary |
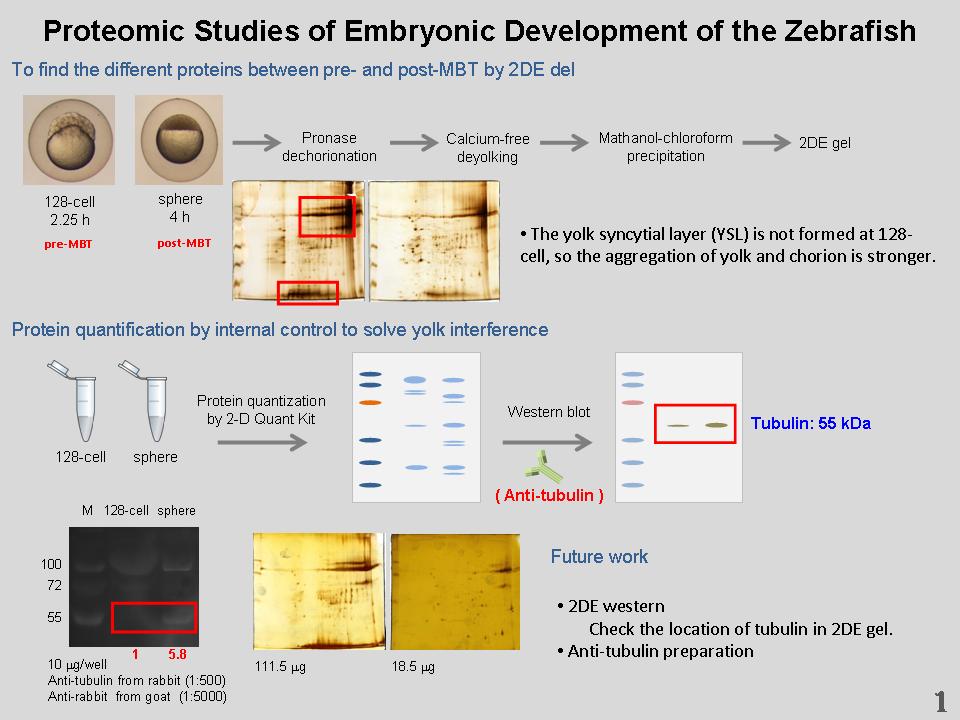
•
1) Yolk syncytial layer (YSL) 的形成機制為何?對於 deyolking 的效果影響有多少? 2) Developmental arrest 是否就是發育阻障 (developmental block)? 3) 由於 128-cell 樣本少且 deyolking 效果不佳,未來將取 256-cell 的樣本。 4) 利用銀染 kit 染色的蛋白質點將定其身份,確認 kit 效果及可能有差異的蛋白質點。 5) 為了避免 yolk 在兩時期的去除效果不一,以 tubulin 進行 Western blot 將此 internal control 定相同量再做 2-DE。 6) 將進行 2-DE 的大片 Western blot,確認 tubulin 在膠上得位置。 7) ECL 呈色結果有問題,可能是抗體存放太久,所以未來將進行 tubulin 的抗體製備。 8) 未來將預測 tubulin 和小鼠之間的差異區間及非後修飾區域,並合成 peptides 免疫小鼠。 9) 若 2-DE 比較效果不佳,也許取兩個蛋白質體直接進行全部蛋白質的 mass 分析,但是工程較為浩大而昂貴。 |
| 10/03/16 |
7 賈 儒 珍 PCS 的各種定點突變 (決定活性分析條件) |
 |
|
| summary |
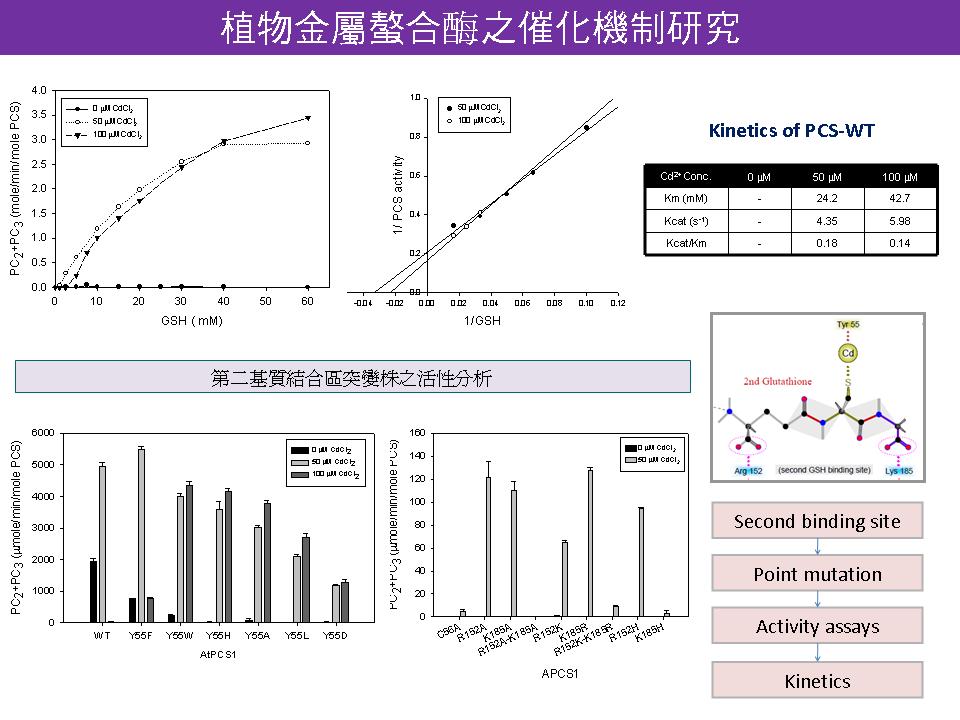
1) 查清楚 E. coli 有無磷酸化系統。 (黃迺茵 follow-up:原核細胞中也有相當的磷酸化現象。) 2) 藻類 PCS 在 T49 附近沒有磷酸化訊號,不能被磷酸化修飾,因此和阿拉伯芥 PCS 相比較,合成 PC 的能力很低。 3) 活性分析結果看起來 50 mM 鎘和 100 mM 鎘處理的 PCS 活性差異不大,用 50 mM 即具有代表性。 4) 活性分析的酵素用量改為 1 mg PCS 較為合理,所有的突變株都用新條件重做一次。 5) 活性分析的條件 (比如酵素定量、鎘濃度和反應時間) 必需可信,才有討論實驗結果的意義。 6) PCS-N 的活性分析條件和其 pI 值過於接近,要調整 pH,先測一次酵素反應的最適 pH 值。 7) 做 kinetics 時,mutants 的反應條件要和 WT 完全一樣,不能有差異。 8) 以 S-methylglutathion 為基質做 kinetics,試試看不能與鎘結合的基質與 PCS 的親和力。 9) C-domain 除了已經做好的 mutation 之外,還需確定是否有其他可能與鎘結合的 Cys 位置。 10) 多重覆幾次 C-domain 的活性分析實驗,確定有再現性後再深入追踨有意義的 mutants。 11) 確定以 second binding site 和 C-domain 之作用機制為論文主題,進行深入的探討。同時也要釐清 PCS 是否在植物中有磷酸化的機制。 |
| 10/03/09 |
11 林 怡 岑 SP-DPE 複合物的對 G3 與 G4 的活性上升 |
 |
|
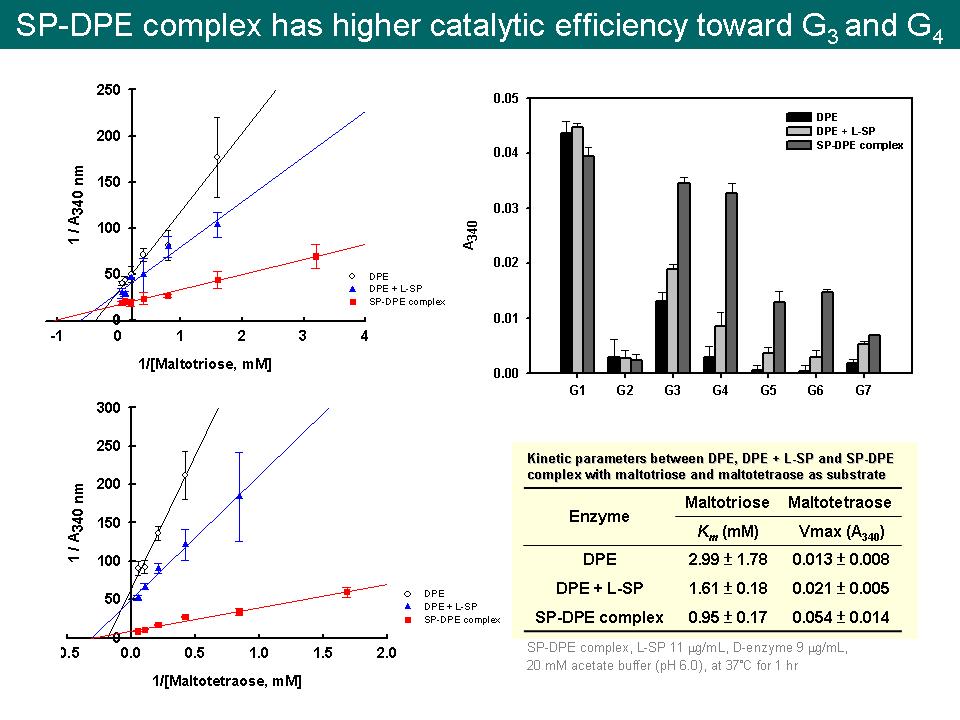
| summary |
實驗結果: 1) 實驗結果顯示 L-SP 與 D-enzyme 結合形成 protein complex,所形成的 SP-DPE complex 對 G3 的親和力增加兩倍,對 G4 的 Vmax 也顯著增加兩倍以上。 建議事項: 1) 綠藻中 PhoB 跟其他型 SP 有何不同? Ans:綠藻內共有 PhoA、PhoB 和 PhoC 三種形式。PhoA 類似高等植物的 L-SP,PhoC 類似高等植物的 H-SP,PhoB 與 PhoA 同樣位於質體中,但 PhoA 對 malto-oligosaccharides 的親和力高,而 PhoB 則對 malto-oligosaccharides 的親和力低。 2) L-SP PI activity 的第一個轉折點產物為何 (從 slow initiation phase 到 rapid elongation phase)? Ans:由王維德 (2006) 與林之儀 (2009) 的實驗結果推得 L-SP PI activity 的第一個轉折點產物為 maltose。 3) 類似 GST-pull down assay 重要實驗的條件必須仔細說明清楚,例如蛋白質 loading 的量、各實驗組的差異,以及 elute 下來的蛋白質量那麼少是否具有可信度。 4) GST-pull down assay 的結果顯示,純化的 L-SP 無法與 GST-DPE 結合,但 crude extract 中的 L-SP 卻可與 GST-DPE 結合,這結果告訴我們什麼? Ans:這代表 crude extract 中的 L-SP 可能有某種後修飾,所以能與 GST-DPE 結合,而純化後的 L-SP,其後修飾可能被破壞了,因而無法與 GST-DPE 結合。L78 有沒有可能扮演某種角色? 5) Confocal 實驗中控制組是如何進行? 為何組織位置並不相同? Ans:控制組所有實驗條件都一樣,但一抗改為 pre-immune serum;組織位置不相同是由於當連續切片放大到 1000 倍後,很難找到完全相同的位置。 6) SP-DPE complex 就是所謂的 HX,但為了與 proteasome 那部份做區隔,D-enzyme 這部份改用 SP-DPE complex。 7) Kinetics 須多重複幾次,更換不同批次的純化酵素進行相同的實驗,假如結果還是確定 Km 或 Vmax 值不會改變的話,可略做調整,使雙倒數圖交於 X 軸或 Y 軸,但不可違背良心判斷。 8) 如何將 SP-DPE complex 與純化的 SP 及 GST-DPE 定相同酵素量進行 kinetics? Ans:目前採用的方法是利用 ECL Western blotting 將目標蛋白質微調到相同的酵素量。 9) Kinetics 的酵素量一定要一樣。 利用 ECL western blotting 定量蛋白質是否可信? 有沒有辦法只利用 SP-DPE complex 本身互相比較?查查看其他 paper 是如何以 complex 進行 kinetics 的? Ans:Tetlow et al. (2008) Analysis of protein complexes in wheat amyloplasts reveals functional interactions among starch biosynthetic enzymes. Plant Physiol. Vol. 146:1878-1891 以 immunoblotting 定量不同來源 protein complex 進行實驗。 10) HPLC 中的 G4~G6 的 peak 都太小,應該尋找最適合的基質濃度 (10 倍 Km) 與反應時間進行實驗,使 peak 更明顯;另外,若要觀察反應的初始產物,剛開始的時間點應該測密集一點。 11) Modeling of SP-DPE complex structure 是用什麼軟體預測的? Protein 接觸面積似乎很小? 瑩貞的 data 顯示 SP 和 DPE 的結合比例是多少? Ans:這是用 Discovery Studio 之 Protein-Protein Docking 軟體預測,另外由瑩貞的 data 顯示 SP 和 DPE 的單元體結合率為 1:1,所以應該可以用 SP 和 DPE 的單元體進行初步預測。 12) 為何 SP-DPE complex 中,若 D-enzyme 將基質傳遞給 L-SP,則初始產物 [G5] ≒ [G4];若 L-SP 將基質傳遞給 D-enzyme,則初始產物 [G4] ≒ [G6]? 13) 為何 SP-DPE complex 的基質傳遞一定是單一方向,而非 shuttle 式的可逆方向? Ans:目前還未看到有 paper 提出 complex 中的 substrate tunnel 為可逆的,但不排除 shuttle 可逆式之可能。 14) SP-DPE complex kinetic 實驗的控制組,除了要有 mixture of L-SP and D-enzyme,還要有 L-SP only 以及 D-enzyme only。 15) Glucan trimming model 中所 trimming 後的短鏈糖大約有多長? Ans:由 Mouille et al. (1996) Preamylopectin processing: A mandatory step for starch biosynthesis in plants. The Plant Cell 8: 1353-1366 結果推測 debranching enzyme 所 trimming 下來的短鏈糖大小約在 G4~G12 之間。 16) Starch synthase 最短所需的 primer 為多長? 會不會真正植物生理並不需特別合成 primer,由 SP-DPE complex 所產生的短鏈糖 (G4 or G5),就可用來當 primer。 Ans:Szydlowski et al. (2009) Starch granule initiation in Arabidopsis requires the presence of either class IV or class III starch synthases. The Plant Cell 21:2443-2457 之結果顯示 SS IV 所需的最短 primer 為 maltotriose (G3)。 17) 澱粉是如何充實的? 需了解澱粉的構造與澱粉合成機制,以及一開始形成澱粉的起始點 (十字中心) 位於組織中的哪部份。 18) 由組織免疫螢光染色的結果顯示 L-SP 和 D-enzyme 好像分布於整個 starch granules 中,並沒有特別集中在某處。 19) 整個實驗架構必須說明一個完整的故事,所以目前的故事是如何? Ans:目前推測 L-SP 和 D-enzyme 所形成的 SP-DPE complex,是先由 complex 中的 L-SP 磷解 G4~G12 (glucan trimming下的短鏈糖),接著再將無法繼續磷解反應的 G4 交由 complex 中的 D-enzyme 進行延長,這樣能有效幫助 L-SP 磷解 MOS 產生 Glc-1-P,供澱粉合成或其他生理反應使用。 20) L78 的斷裂與否跟這整件事情是否有關,需進一步探討。 21) Szydlowski et al. (2009) Starch granule initiation in Arabidopsis requires the presence of either class IV or class III starch synthases. The Plant Cell 21:2443-2457 之結果顯示在 ss III-ss IV double mutant 中,L-SP 的活性提高了接近 10 倍,這代表什麼生理意義? 未來工作: 1) BiFC 可同時利用 sweet potato roots 和 rice 兩種物種的 L-SP 與 D-enzyme 分別進行。 2) 設計不同的 kinetics 實驗 (改變酵素、基質和時間),觀察酵素的 vo、Km 與 Kcat,推導出代謝方向,由簡單的實驗可解決重要的問題。 |
| 10/02/09 |
13 何 杰 龍 主題 |
|
slide |
|
| summary |
|
| 10/01/19 |
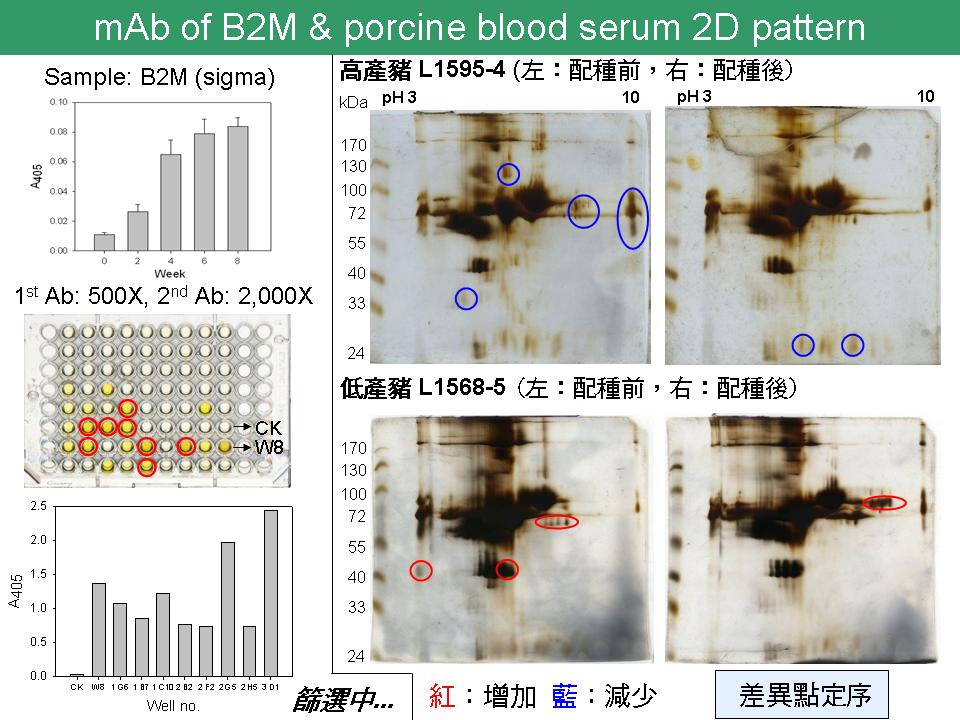
2 沈 志 昱 尋找豬胚胎發育的 marker |
 |
|
| summary |
Part 1 1) B2M 在何時期多、何時期少? MHC protein 在一般細胞都存在,是否已知在胚胎中的意義? 與林老師討論選此三目標的原因。為何選 IGF 而不選其他如 EGF、FGF 等。 2) B2M 有 3D structure 則可確認 peptide 設計之位置。 3) 挑 master cell 時亦可找 ELISA 吸光值較低的,以增加抗體辨認位置之多樣性。 4) 可用 albumin 抗體確認是否 albumin 去除乾淨,可購買或自製。 5) 用抗體偵測出的差異點,才是 2-DE 上真正有意義的差異 (但還是要有 CBR 染色比較)。 Part 2 1) Lane & Gardner 在 2005 年於 JBC 的報告,是否提及 malate-aspartate shuttle (MAS) 在 embryogenesis 中的角色?若 MAS 十分重要,也許可以當 marker,可從代謝產物 (metabolomics) 或酵素著手,嘗試對此系統做抗體。 [代謝圖] 2) 在胚胎發育過程,LDH 的含量逐漸遞減。2-cell 期時因 MAS 剛啟動,需較大量 LDH 將 lactate 轉換成 pyruvate 產能,LDH 與 MAS 似乎有互相牽制的密切關係,但 lactate 從何而來?此外,8-cell 期胚胎開始利用 glucose 產能,要如何解釋此時 LDH 與 MAS 的關聯?亦或此時 MAS 轉為與其他蛋白質互相牽制? 3) 從 1-cell 至 8-cell,胚胎產能利用的原料不同,各時期中 pyruvate / lactate 的轉換,與 NAD+/NADH 的轉換,是否能更清楚描繪可能的機制? 此外,試著了解 glycolysis 與 PPP 的相互關係。 |
| 10/01/01 |
0 報 告 者 主題 |
|
slide |
|
| summary |
|
{kind=link}