研 究 主 題
The structure and function of starch phosphorylase from sweet potato roots
▼ 磷解脢構造.活性調節.降解過程.可能機制.生理角色.研究題目.References ▲ 重金屬研究
|
研 究 主 題 |
||
| 甘藷澱粉磷解脢活性調節之分子機制 | ||
|
The structure and function of starch phosphorylase from sweet potato roots |
||
|
|
||
|
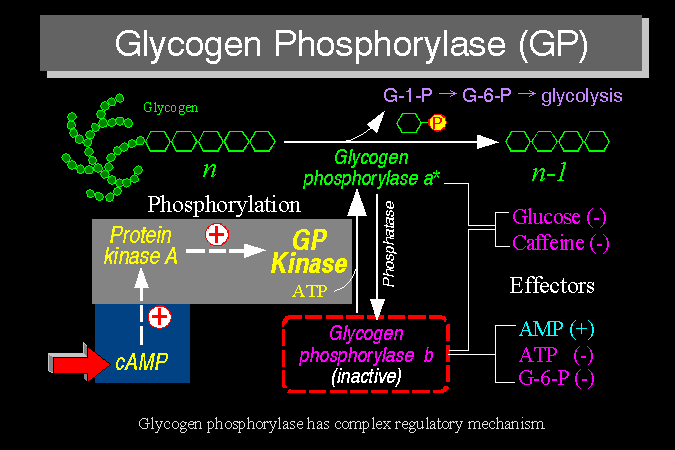
1) 動物的 肝糖磷解脢 (glycogen phosphorylase, GP) 影響動物體內血醣濃度高低,有很複雜的酵素調節機制,例如:Ser-14 的磷酸化及 allosteric effect 等。 植物的 澱粉磷解脢 (starch phosphorylase, SP) 則是 GP 在植物細胞的對等酵素,可分解植物的貯藏性多醣,但卻缺乏類似上述 GP 的複雜酵素調控系統。下圖是 GP 的活性調節機制摘要。 |
||
|
|
||
|
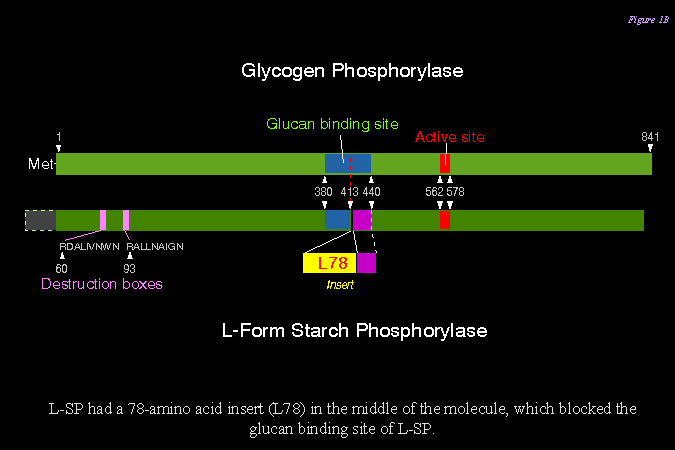
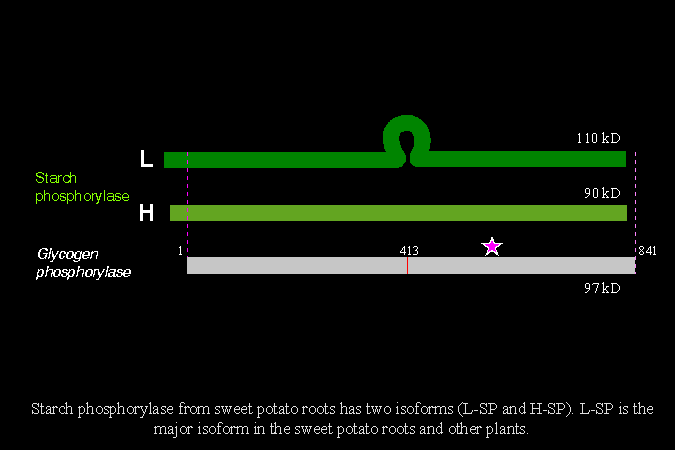
2) 比較這兩者的基因序列及分子構造 (下圖),發現甘藷 SP 分子中央,多出一段由 78 個胺基酸組成的片段 (L78),剛好阻礙 SP 對基質的結合區 (glucan binding site),在立體構造上會阻礙 SP 與其基質的結合。 |
▲ TOP |
|
|
|
||
|
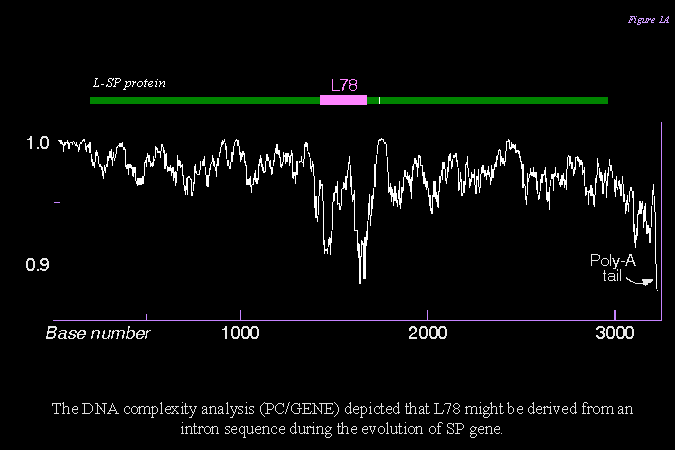
由 SP cDNA 的序列複雜度分析結果推斷,L78 可能 是由 intron 所衍生而來的。 |
||
|
|
||
|
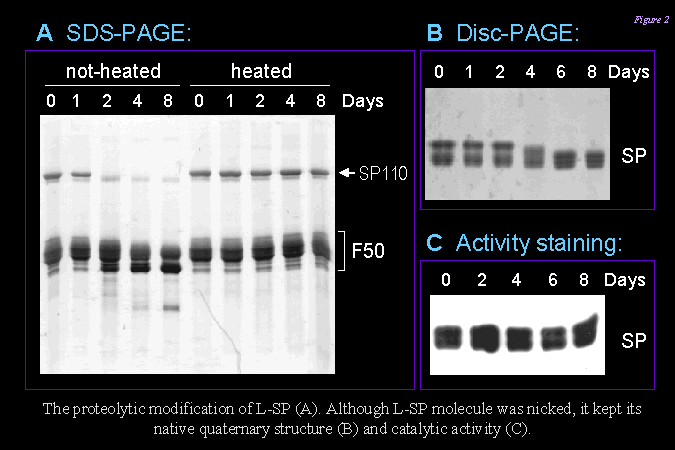
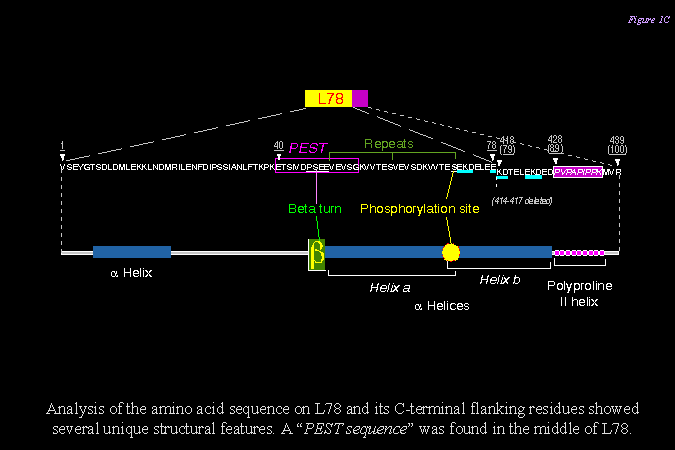
3) 而 L78 上具有 PEST 序列,分子容易受到降解,因此原態 SP (110 kD x 2) 在甘藷貯藏過程中很容易斷裂,在 SDS 電泳上可看到一群分子量在 50 kD 左右的片段 (下圖 F50)。 請注意 SP 分子雖然容易從中間斷裂,但仍然保持其原態分子構型 (下圖 B) 及其酵素活性 (下圖 C)。 |
||
|
|
||
|
4) 上述 SP 因為有 L78 存在,阻礙了酵素與澱粉的結合,因此與澱粉的親和力較小,稱為 L 型 (low affinity, L-SP);植物體中另有一種與澱粉親和力較大者,稱為 H 型 (high affinity, H-SP),其分子內則缺乏 L78 片段。 |
||
|
|
▲ TOP |
|
|
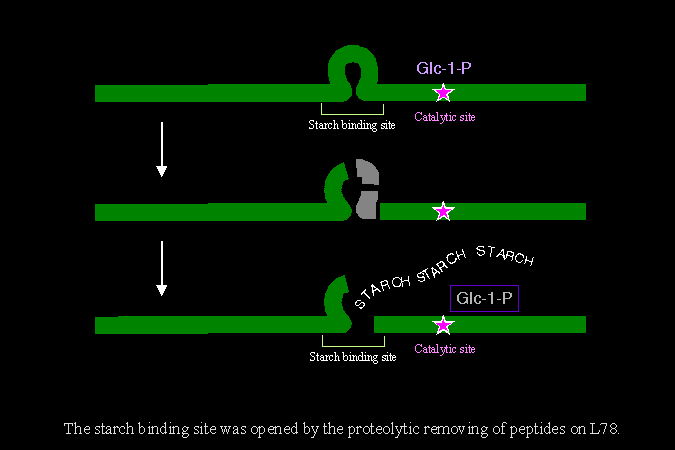
1) 基於以上觀察,植物的 SP 可能並非完全沒有調節機制,SP 可能先以類似前驅物的方式存在,此時酵素分子因為有 L78 的立體障礙,對澱粉的結合力很低;當 L78 被蛋白脢切開之後,活性才提高起來。 |
||
|
|
||
|
2) 雖然 H-SP 的比活性較高,但相對於 L-SP 則其含量很低,同時在不同組織的分佈也有差異;因此,H 與 L 型 SP 的消長與分佈,可能也是其調節機制之一種方式。 |
||
|
|
||
|
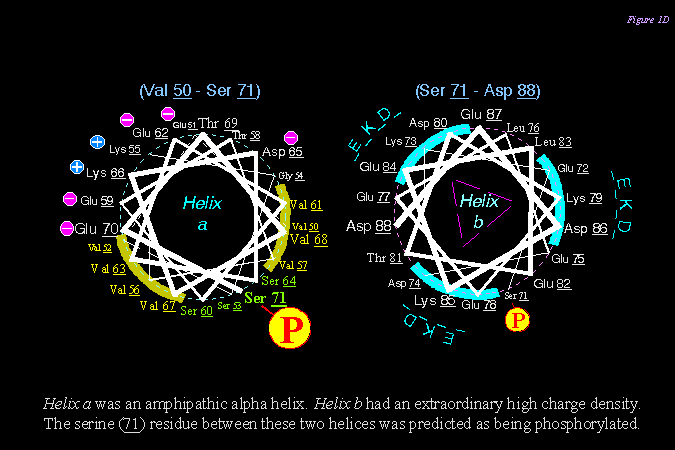
3) 下圖說明 L- SP 分子上 L78 的胺基酸序列,發現其二級構造有許多有趣的特徵,可能與其降解有關 (PEST site, 磷酸化基團, polyproline II helix 等)。 |
||
|
|
|
|
|
L78 的後半段捲繞成 a helix,其構造也非常特別 (下圖),可能有特殊的生理功能。 |
||
|
|
▲ TOP |
|
|
|
|
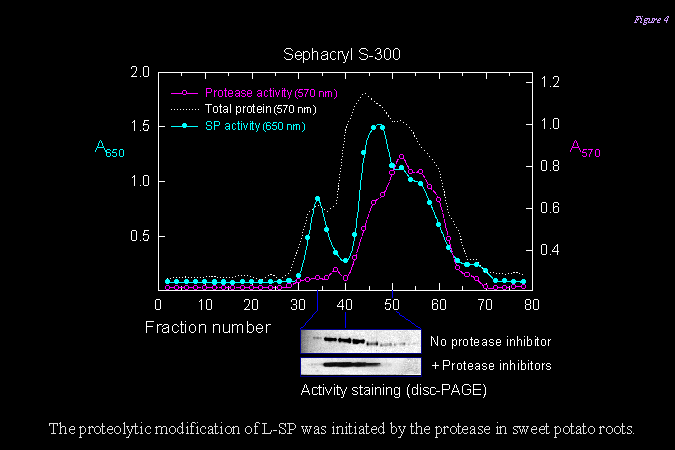
1) 以膠體過濾管柱純化 SP 酵素,發現其活性分劃涵蓋範圍很廣 (下圖),在 disc-PAGE 上的泳動圖譜也有微細的變化,暗示著 SP 分子在塊根的生理發展過程中,進行著一連串的修飾與改變。而此種改變,可以被 protease inhibitor 所抑制,因此是一種蛋白脢水解的修飾。 |
||
|
|
||
|
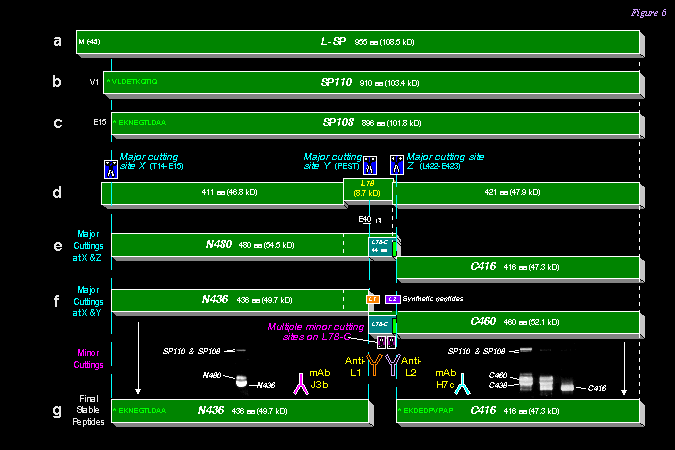
2) 取 SP 進行部份胺基酸序列分析,發現其分子是斷裂在 L78 的末端;也就是說原先由 intron 所表現出來的部份,最後可能完全被切除。我們已推出 L-SP 在 L78 上的降解程序 (下圖),並以抗體檢定所產生的大小片段。 |
||
|
|
||
|
3) 另發現一個新的切斷點,是在 N-端第 15 胺基酸,而在 GP 分子上正是可供磷酸化的 Ser-14 處,這可能也是用來調節 SP 活性的方法。 |
||
|
|
||
|
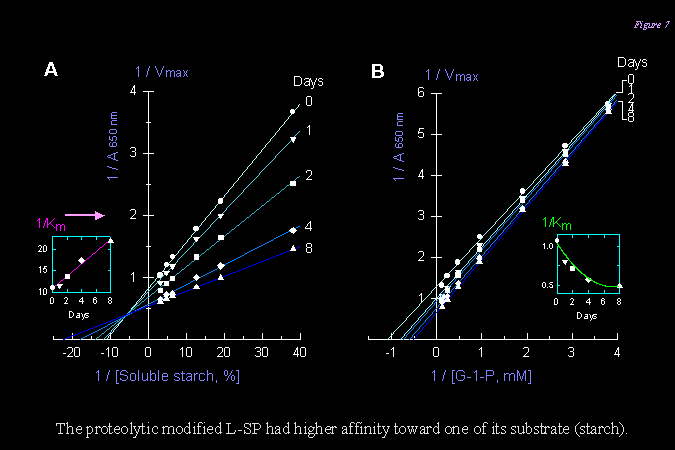
4) 在 SP 貯藏一週間,完整的 110 kD SP 分子逐漸消失,同時觀察到 SP 比活性增加 (約 20%);在此過程以動力學觀察澱粉合成方向的反應,發現 SP 對澱粉的 Km 下降,而 Vmax上升 (如下圖)。 |
||
|
|
▲ TOP |
|
|
四、澱粉磷解脢可能以 ubiquitin-proteasome 程序降解: |
||
|
1) 在 SP N-端附近序列,發現有兩個典型 destruction boxes,顯示 SP 分子可能進行 ubiquitination,而蛋白質 ubiquitination 的結果之一,就是被 proteasome 降解。 我們已在甘藷塊根中,發現有相當多量的 ubiquitin 及 proteasome。 |
||
|
|
||
|
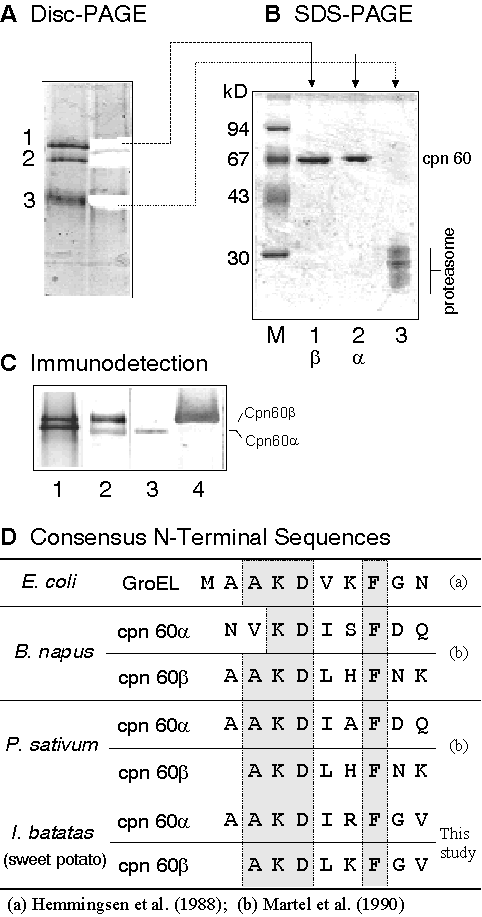
2) 在 SP 的純化過程中,一直有高分子量蛋白質伴隨,經抗體檢定後發現是 hsp60,推論 SP 可能需要 chaperone 的輔助;可能是幫助 SP 進入造粉體,或幫助降解後的分子維持原態。下圖是分離及檢定甘藷塊根中 proteasome 及 chaperone 的過程。 |
||
|
|
▲ TOP |
|
|
|
|
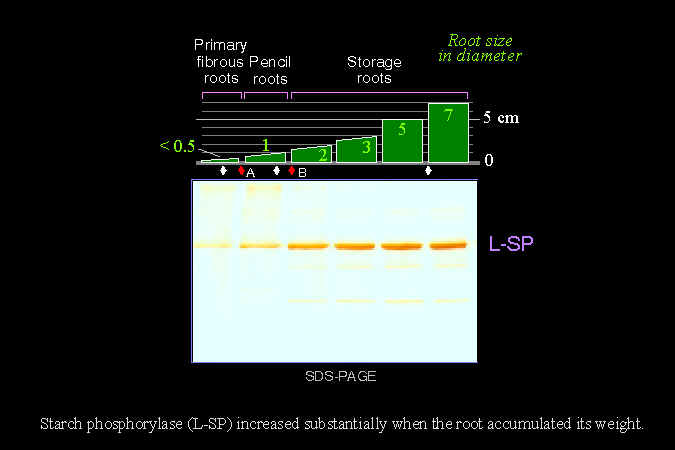
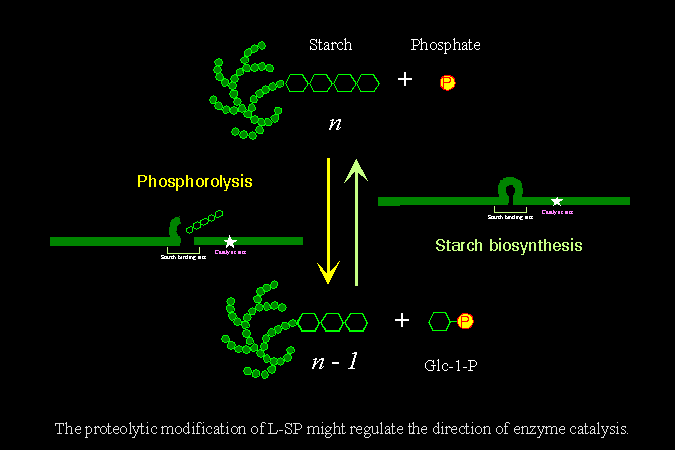
1) 目前仍不確知澱粉磷解脢的真正生理角色。 雖然一般均認為 SP 是用來磷解澱粉,產生磷酸葡萄糖;但我們在試管中發現,顆粒性澱粉粒並非 SP 的良好基質。同時,很多實驗都證實,L-SP 的存在與植物澱粉的累積量成正比。 |
||
|
|
||
|
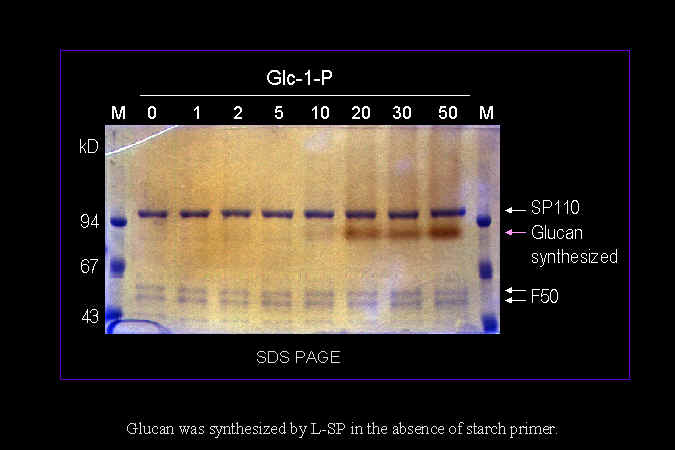
2) 我們確定 L-SP 可以使用 Glc-1-P 作為唯一的基質,在試管中合成長鏈澱粉分子。 |
||
|
|
||
|
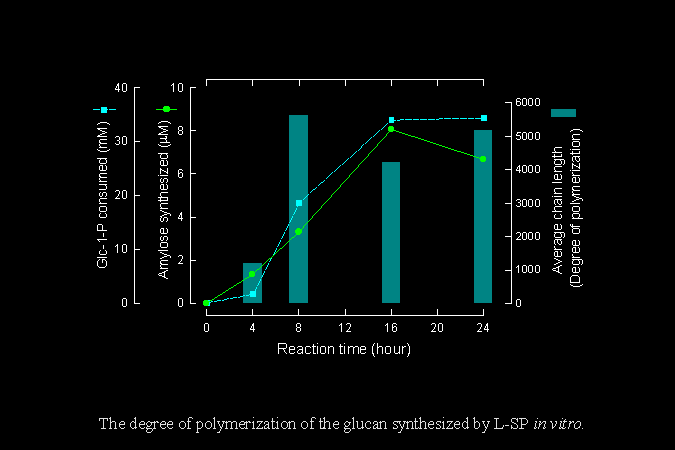
測定由 L-SP 合成出來的澱粉分子,約含有 4,000~6,000 個葡萄糖單位。 |
||
|
|
||
|
3) 有關植物細胞澱粉粒的生合成途徑,其 澱粉引子 (primer) 的來源,目前仍無定論。 我們由以上觀察提出一個假說,認為 L-SP 可能是合成澱粉引子的酵素,而 L-SP 分子的活性又受到各種調節因素的控制 (可能在 L78 上),以便引發細胞內澱粉合成的開始。 |
||
|
|
▲ TOP |
|
|
|
|
References (protein phosphorylation and degradation) |
||
|
Hochstrasser M (1996) Protein degradation or regulation: Ub the judge. Cell 84: 813-815 |
||
|
St-Pierre B, Bertrand C, Camirand A, Cappadocia,M, Brisson N (1996) The starch phosphorylase gene is subjected to different modes of regulation in starch-containing tissues of potato. Plant Mol Biol 30: 1087-1098 |
||
|
Verslues PE, Braun DM, Garcia MXU, Stone JM (1996) Protein phosphorylation: Examining the plant CPU. Trends Plant Science 1: 289-291 |
||
|
Goldberg AL, Elledge SJ, Harper JW (2001) The cellular chamber of doom (cellular structures call proteasomes recycle old proteins). Scientific American 284(1): 56-61 |
||
|
References (Juang, RH) |
||
|
Chan HY, Lin TY, Juang RH, Ting IN, Sung HY, Su JC (1990) Sucrose synthetase in rice plants: growth associated tissue specific distributions. Plant Physiol 94: 1456-1461 |
||
|
Chang SC, Lin PC, Chen HM, Wu JS, Juang RH (2000) The isolation and characterization of chaperonin 60 from sweet potato roots. Involvement of chaperonins in starch biosynthesis. Bot Bulletin Acad Sinica 41: 105-111 |
||
|
Chen HM, Chang SC, Wu CC, Cuo TS, Wu JS, Juang RH (2002) The catalytic behavior of L-form starch phosphorylase from sweet potato roots is regulated by proteolysis. Physiol. Plantarum 114: 506~515 |
||
|
Chern MS, Mo YC, Juang RH, Su JC (1990) Probing the protein structure of sweet potato starch phosphorylase with monoclonal antibodies. J Chinese Biochem Society 19: 55-64 |
||
|
Chiang CL, Lu YL, Juang RH, Lee PD, Su JC (1991) Native and Degraded Forms of Sweet Potato Starch Phosphorylase. Agric Biol Chem 55: 641-646 |
||
|
Young GH, Chen HM, Lin CT, Tseng KC, Wu JS, Juang RH (2005) Site-specific phosphorylation of L-form starch phosphorylase by a protein kinase activity from sweet potato roots. Planta (in press, published on line) |
||
|
Linkings |
||
|
Boston Biochem (以 ubiquitin-proteasome 為主的供應廠商) |
||
|
Ubiquitin and the Biology of the Cell (1998 年出版的大全集, Plenum; check Amazon) |
||
 |
||
| 建立日期:2000/4/13; 更新日期: 2010/05/29 | ||
|
本網頁資料隨時在編修中 © 版權所有 |
||
|
▲ TOP |
||
▲ 磷解脢構造.活性調節.降解過程.可能機制.生理角色.研究題目.References ▼ 重金屬研究