|
2008 進度報告摘要 |
|
| 08/12/24 |
4 賈 儒 珍 L-SP 及各種突變的蛋白質表現 |
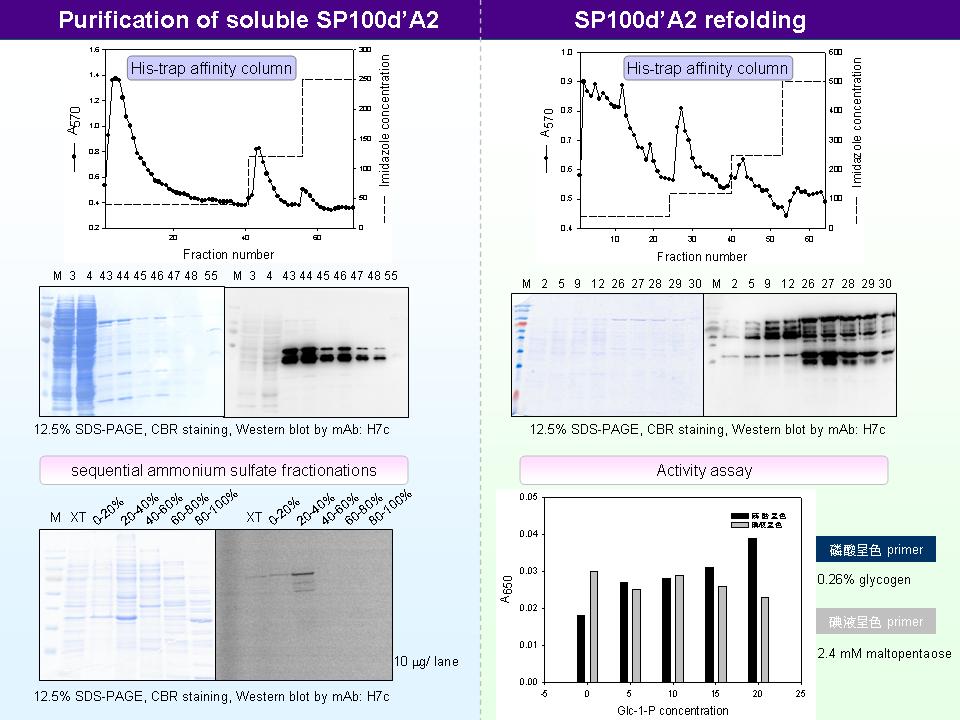 |
|
| summary |
1. 目前正在進行 L-SP 全長和真核表現載體的 cloning,未來將嘗試在昆蟲細胞表現。 2. 已經在 SP100d’ 的 N-端和 C-端中間插入一個 TEV protease 切位,之後表現出 SP100d’ 可以在試管內先用 protease 處理,再做 L78 rescue 實驗。此種作法可以鬆開 starch binding site,並減少 L78 docking site 被遮蔽的疑慮。 3. 因為 SP 上有很多 rare codons,無法一一修正,在大量表現時會有多個 alternative initiation site,使得全長被轉譯成多個片段。已經把兩個確定 alternative initiation sites,從 Met 突變成 Ala,並把此突變株命名為 100d’A2;但是純化出的表現蛋白仍有很多較小片段。 4. 用 His-trap affinity column 純化出的表現蛋白,全長的比例遠低於粗抽的量,可能是全長的可溶性較低,或是 his-tag 無法抓住 100 kD 的完整表現蛋白。 5. GST tag 用於表現蛋白的純化效果很好,但是考慮到 GST 的大小 (約 20 kD) 而不使用。可以嘗試使用大小約 7-9 個胺基酸的 HA tag 和 Flag tag。 6. 用不同硫酸銨分劃部分純化 SP100d’A2 表現蛋白,表現蛋白主要在 20-40% 的硫酸銨飽和濃度下被沉澱出。但是在 20-40%、40-60%、60-80% 和 80-100% 飽和濃度,都有澱粉磷解酶的活性。應該要做 mock 來比較活染結果,以確是內生性蛋白質或是表現蛋白的活性。 7. 在有 PLP 的環境之下做 SP100d’A2 的 refolding,希望能夠藉由 PLP 幫助並穩定表現蛋白的摺疊。Refolding 後得到的表現蛋白活性仍然很低,應改變基質濃度、反應時間等因素,並測試磷解方向反應的活性。 8. 需做 native PAGE 或 gel filtration,確定純化後的表現蛋白四級構造是 monomer 或 dimer。 9. 可以在跑完 native PAGE 之後外加 L78 和 Glc-1-P 做 in gel rescue 的實驗,利用碘液染色就可以測得膠內的產物。 10. 測試表現蛋白活性的基質,使用澱粉會比肝醣好 (畢竟還是 truncated L-SP)。 11.因為無法取得甘藷 L-SP 的突變株,因此將嘗試使用阿拉伯芥的 Atphs1 突變株做表型分析。阿拉伯芥種子中的儲藏性澱粉和葉子中的澱粉特性可能不同。 12.可以買到兩種 Atphs1 突變株,其中 Atphs1-2 已知 T-DNA 插在 5’-UTR 中。 13.突變掉 alternative initiation site (Met→Ala) 也許會影響蛋白質功能,利用真核表現系統可以完全避免 rare codons 問題。
|
| 08/12/10 |
11 何 杰 龍 禽流感病毒外鞘蛋白 HA1 上醣類部份的探討 |
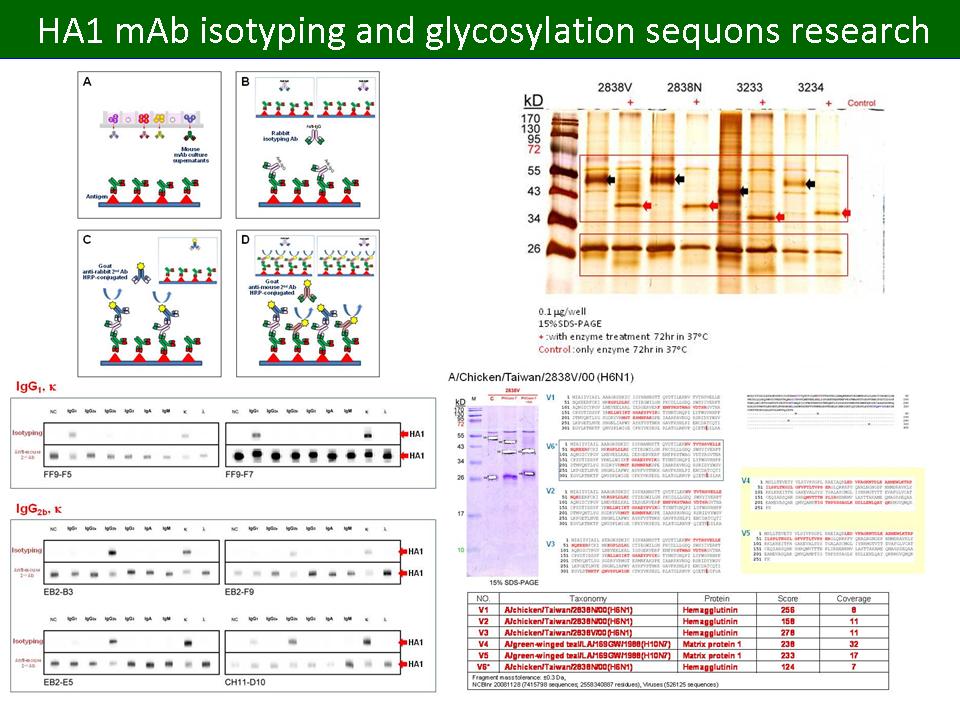 |
|
| summary |
1. AIV mAb bank 目前已篩出 6 株對 H6 亞型具專一性之 HA1 單株抗體,效價高達 16,000 x 以上,因為這些抗體的免疫呈色圖譜非常類似,我們一度認為可能全是同一株。 2. 進一步進行 antigen-dependent mAb isotyping,發現這 6 株 HA1 單株抗體可分為 IgG1 與 IgG2b 兩類。據此推測:2838V 病毒的 HA 上有一些較強的抗原決定基 (epitope),因而各種抗體多辨認這些位置。 3. 以 PNGase F 切除 N-link 的醣基後,2838V 與 2838N 病毒株的 HA1 由 55 kD 降至 38 kD 左右,與推測的 HA1 無修飾胺基酸序列片段 (348 aa) 分子量相符。 4. 以 LC/MS/MS 檢測 PNGase F 處理後,AIV 蛋白質 SDS-PAGE 電泳圖譜 55 kD 與 38 kD 的條帶,皆為 HA1。 5. 繼續進行 AIV 蛋白質醣基化位置對蛋白質體及致病機制影響的研究。 6. 搜尋目前醣基化位置影響 AIV 致病機制的相關文獻。 7. 比較 2838V 與 2838N 的 HA1 序列差異,並與已知結構的病毒株 (H5N1 或 H1N1) 進行比對。 8. 積極進行 in vivo 的測試。 9. 比較 PNGase F 切除 AIV HA 醣基後 2-DE 圖譜之差異。 10. 報告內容要精簡並聚焦於重點。 |
| 08/11/19 |
1 謝 瑩 貞 SP 與 D-enzyme 的交互作用 |
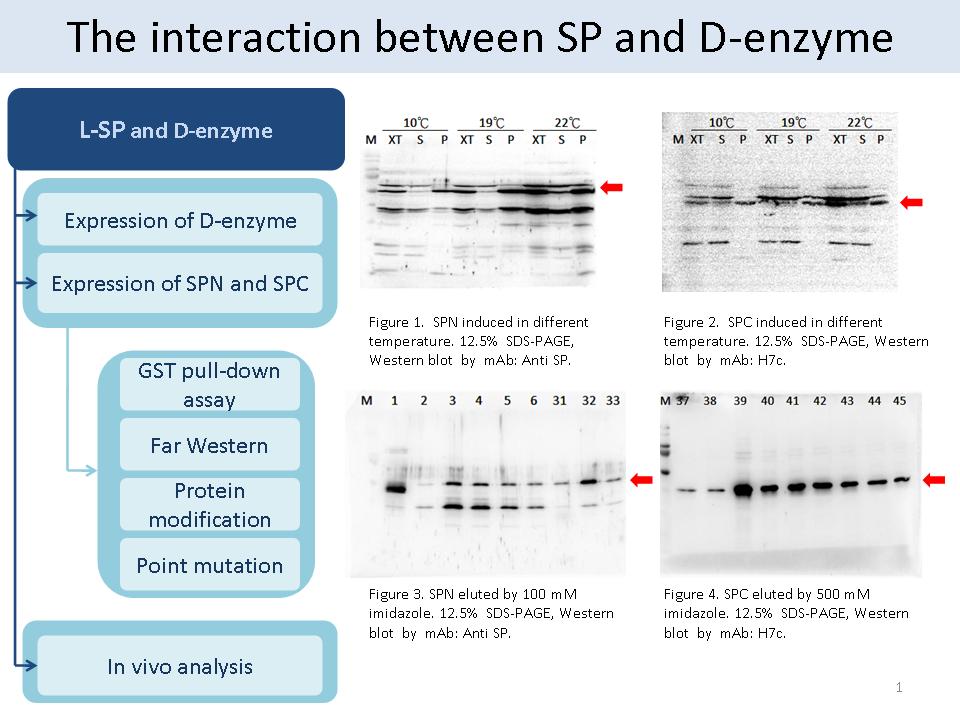 |
|
| summary |
1. 觀察 SP與 DPE 結合之目的? 藉由觀察 SP 與 DPE 之交互作用,找出 DPE 在澱粉代謝過程中所扮演的角色。 2. SPN 與 SPC 的表現量多並不多;D-enzyme 的表現大約為 1 mg/L。 3. SPN 與 SPC 純化後,經 SDS-PAGE 所呈現的分子量是正確的,造成很多 Band 的原因可能是 alternative translation。 4. In vivo 能有相同反應才有實驗價值,若蛋白質只是在打破細胞時才有 binding,則在生物體中可能沒有作用。 5. 比較實驗結果後,完整的 SP110 似乎比 F50 容易與 DPE 結合,也有可能是在 L78 結合。 6. 在 GST pull-down assay 中,bait 為含有 GST 的 D-enzyme ; prey 則是可能會與 D-enzyme 結合的蛋白質。 7. 將 SPN, SPC, L78, SPN+L78, SPC+L78, XT, pure SP 分別去做 GST pull-down assay。 8. 以 DPE 為實驗主題,分別做不同時期的觀察,與 SP 在不同時期的作用是否改變。 9. 可利用 Far Western 觀察蛋白質間的交互作用。 10. 不見得要完全相信 paper 的推論,以不同的觀點去分析結果,才能有新的發現。 11.繪製實驗的流程圖 (合併學姊的實驗流程),並注意文字符號的規則,例如:2D PAGE 要改成 2-DE,每頁 slide 上方要有簡短明確的主題。 |
| 08/11/12 |
1 劉 怡 君 製備 AIV PA, PB 單株抗體 |
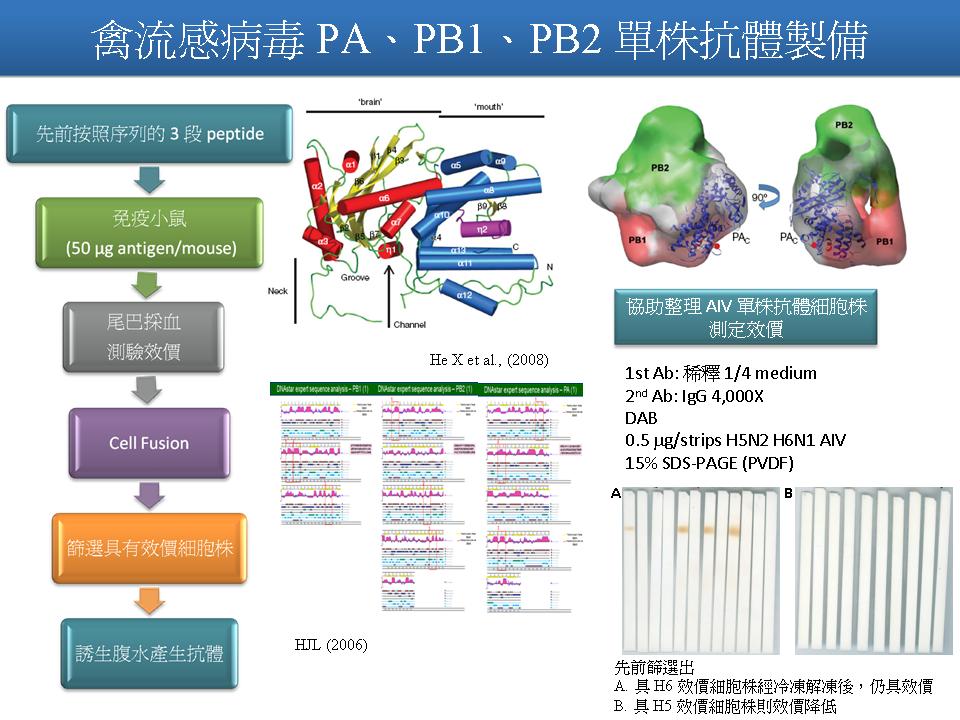 |
|
| summary |
1. 查看 glyceraldehyde-3-phosphate dehydrogenase 與 (-) strain RNA 的關係。 2. 免疫小鼠,若有效價,立刻去染小片 2DE 確定。 3. 是否嘗試表現蛋白? 4. 進一步思考,做出抗體後下一步的研究,以便說完整的故事。 5. 比對此一 RNA dependent RNA polymerase 和真核生物的 RNA polymerase 相似度。 未來工作: 1. 以 peptides 繼續免疫小鼠,嘗試製備 PA, PB1, PB2 的單株抗體。 2. 以抗體定位 PA, PB1, PB2 於 2DE 電泳圖譜位置,與 1978 Martin 結果比較。 3. 協助整理 AIV 單株抗體細胞株,測定效價。 |
| 08/10/29 |
1 黃 迺 茵 AtPCS1 活性區的關鍵胺基酸突變實驗 |
 |
|
| summary |
1. 深入了解進行突變 R152A、R152E 和 K185A 之研究目的。 2. Point mutation 不順利的原因,經剔除掉 DpnI 壞掉或 program 有問題的可能,應為操作技術的問題 (例如 pippeting 不確實、reagents 添加量不準確)。 3. 購入 PCS 突變株種子後 (要說明何種突變株?),應進行 homozygote 篩選,以確定 PCS activity 完全消失。其方法為:突變株種子播種後,第一次自交的種子 (F1) 依照孟德爾遺傳定律,應該有 1/4 為 homozygote,依 phenotype 挑出這些突變株,採收其自交後的種子 (F2) 應有很高機率為 homozygote。 4. 可利用 PEG fusion 將 Ti plasmid 送入 Suspension cell 內,暫時表現基因 (有何目的?)。 5. 要做 complementary mutation 以確定突變株的 PCS activity 是否可因而回復 PCS 基因表現。 |
| 08/10/22 |
3 黃 惠 敏 AtPCS1 蛋白質的純化與結晶條件探索 |
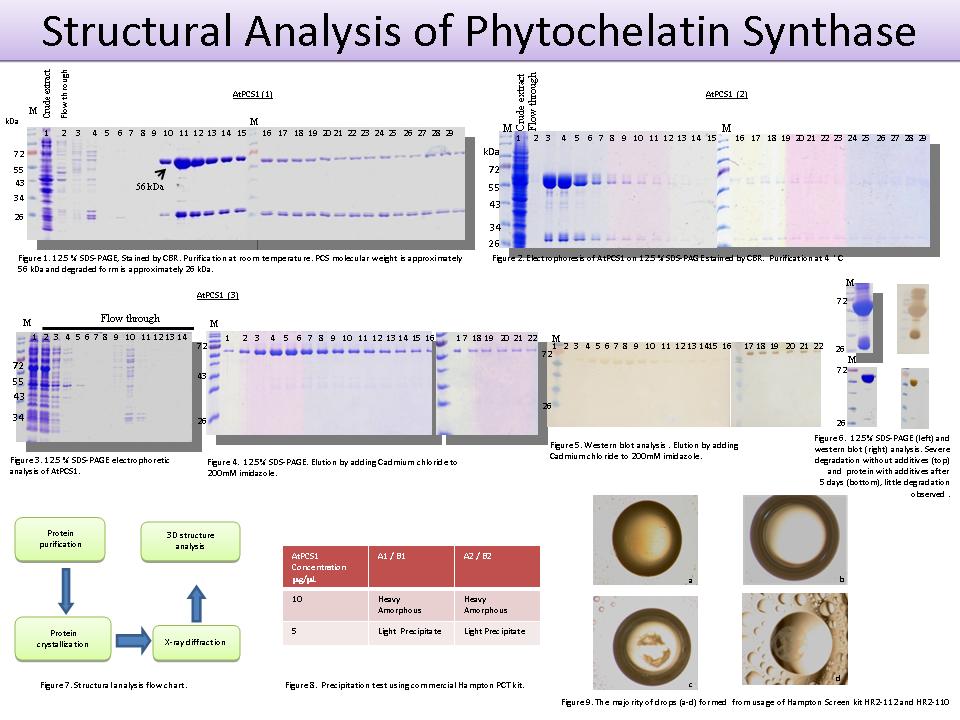 |
|
| summary |
1. AtPCS1 表現蛋白質在純化過程中容易降解。 2. 於 4C 下還是有 60% 之降解。 3. 加入 CdCl2 可以有效減緩降解至 5~10%。 4. 如此的蛋白質可以維持至少五天不致降解。 5. 以 7C5 抗體檢測 AtPCS1 還是有降解現象。 6. 結晶實驗以 1:1 混合後的 蛋白質濃度 5 mg/mL。 7. 然而一週後仍無傑接出現 (Hampton HR2-110 及 HR2-112)。 8. 降低保存緩衝液的 Tris 濃度至 10 mM,並且儘量去除所含的 10% 甘油。 9.至中研院繼續嘗試結晶條件。 |
| 08/10/08 |
3 羅 庭 芳 不同本土禽流感病毒株之蛋白質體與毒性強弱 |
|
|
|
| summary |
1. 使用 2 株本土禽流感病毒 (獸醫系王金和老師) A/chicken/Taiwan/2838V/00 (H6N1);A/chicken/Taiwan/2838N/00 (H6N1)。 2. 2838V 由 2838N 突變而來,HA 突變率 1.41% 共有 8 個突變位,兩者 HA1 peptides 差了三個 charge。 3. 以 CIAP 及 anti-phospho-Ser, anti-phospho-Thr, anti-phospho-Tyr 與 Pro-Q Diamond 得知 HA1 無磷酸化,故 HA1 的 pI 值的差異不是來自於蛋白質磷酸化,而 M1 protein 則有磷酸化。 4. 以 neuraminidase 酶切 sialic acid ,可發現 pI 值較高的 HA1 逐步消失,且在 24 kD 處出現新的蛋白質點,質譜儀定序結果得知 24 kD protein 為 M1 蛋白。 5. Lectin blot 證明 HA1 及 M1 protein 上有 sialic acid,HA1 有 a2-3 linkage 的 sialic acid,M1 protein 則是同時有 a2-3 及 a2-6 linkage 之 sialic acid。 |
| 08/09/24 |
3 林 之 儀 甘藷塊根 L-SP 之 PI 活性催化機制 |
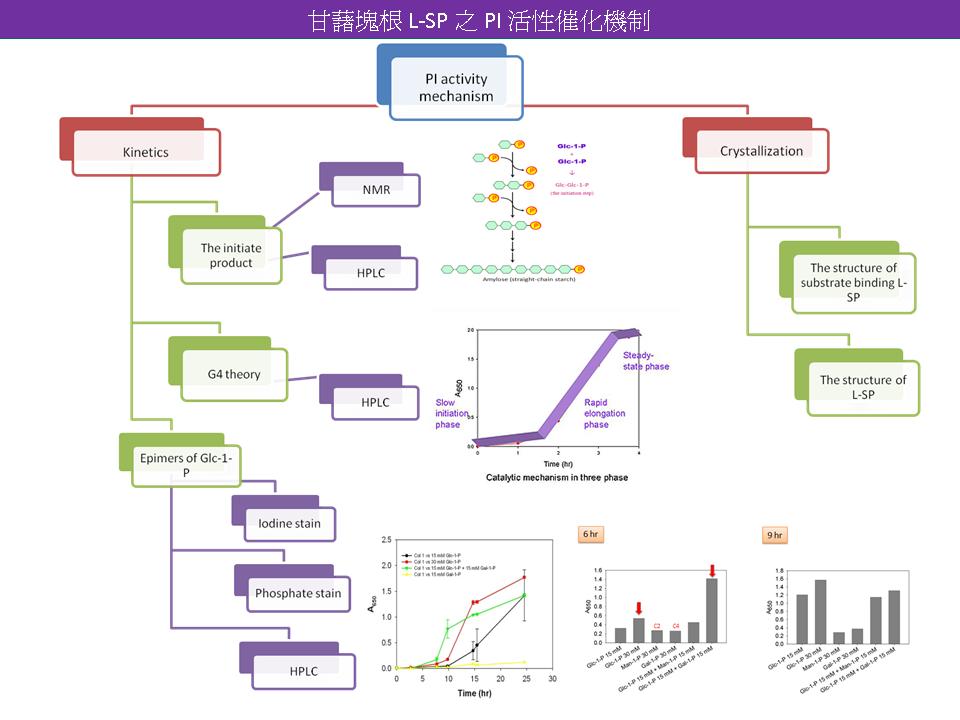 |
|
| summary |
1. 測定 epimer 對 PI activity 影響實驗,為何選用 30 mM Glc-1-P 及 15 mM Glc-1-P? 30 mM Glc-1-P 是否為 SP 的 Km? 應了解設計實驗的原理。 2. Epimer 的 PI activity 所得產物為何?是否可用 MS 或 NMR 圖譜測定醣組成。除了磷酸呈色,需再測定碘液呈色。 3. 養結晶時使用 GP 的 activator AMP 是否適合? SP 會 binding AMP 嗎? 4. PI activity 的最初產物可以經由 HPLC 來確定。 5. G4 theory 是否可偵測產物之醣結構來證明? 6. 需綜合學長姐們的實驗,進行更深入的探討。 Future work 1. 使用碘液呈色來確認 Glc-1-P 的 epimer 之 PI activity 其醣鏈生成是否符合磷酸呈色的結果。 2. 使用其他的 C4 epimer 進行 PI activity 反應,看其是否亦會有類似結果。 3. 以 HPLC 測定 Initiation step 之產物及探討是否 PI activity 從 G4 開始進入 Rapid elongation phase。 4. 使用 NMR 圖譜進行醣結構分析。 5. 繼續進行養晶。 6. H-SP 之純化及酵素動力學分析。 |
| 08/09/17 |
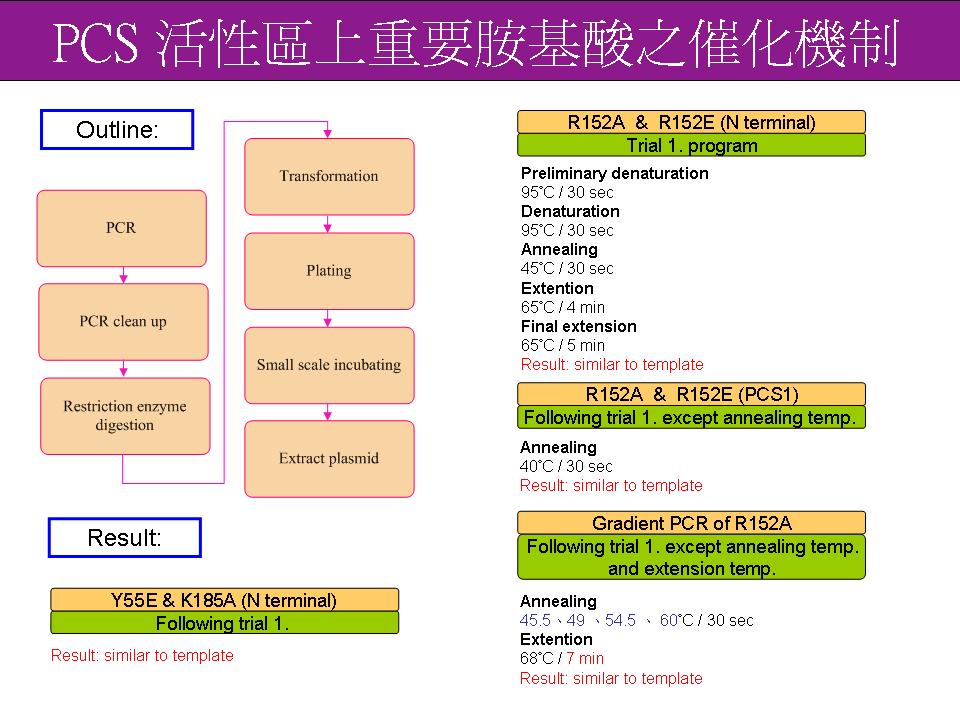
4 眭 毓 庭 PCS 活性區上重要胺基酸之催化機制 |
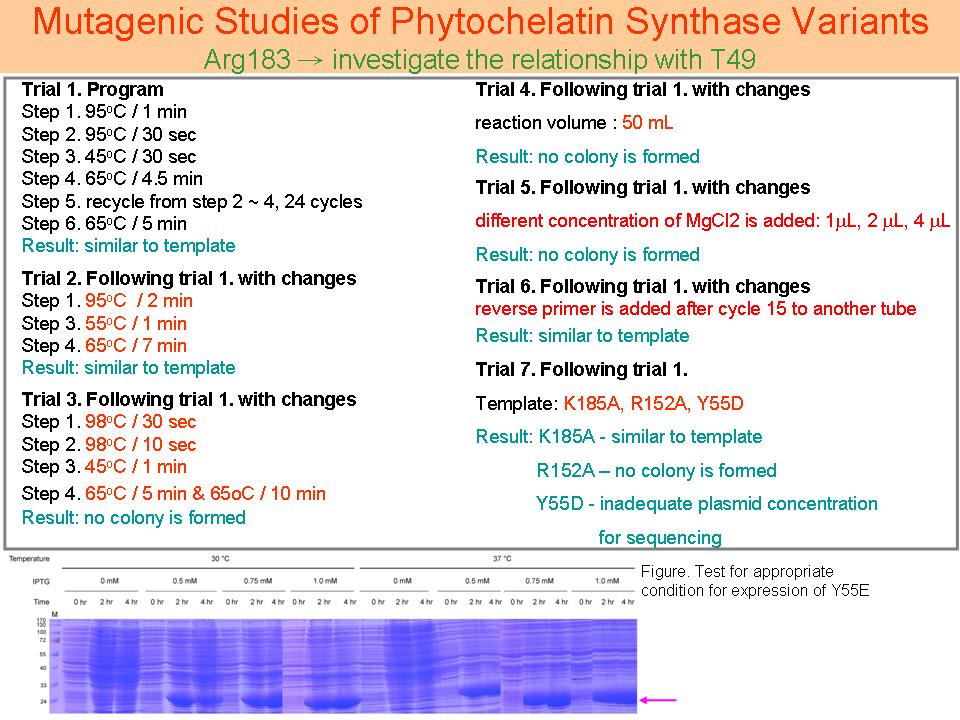 |
|
| summary |
1. 表現條件結果還是必須要以 western blotting 來確定 2. 先集中做 Y55 位置的點突變: Y55E、Y55D、Y55F、Y55W 四種,其他位置如 R183 和 K185 以後再做。 3. 嘗試更改 PCR 條件中 primer / template 比例,找出最適比例進行 PCR。 |
| 08/09/10 |
3 賈 儒 珍 甘藷澱粉磷解脢重組蛋白之表現 |
 |
|
| summary |
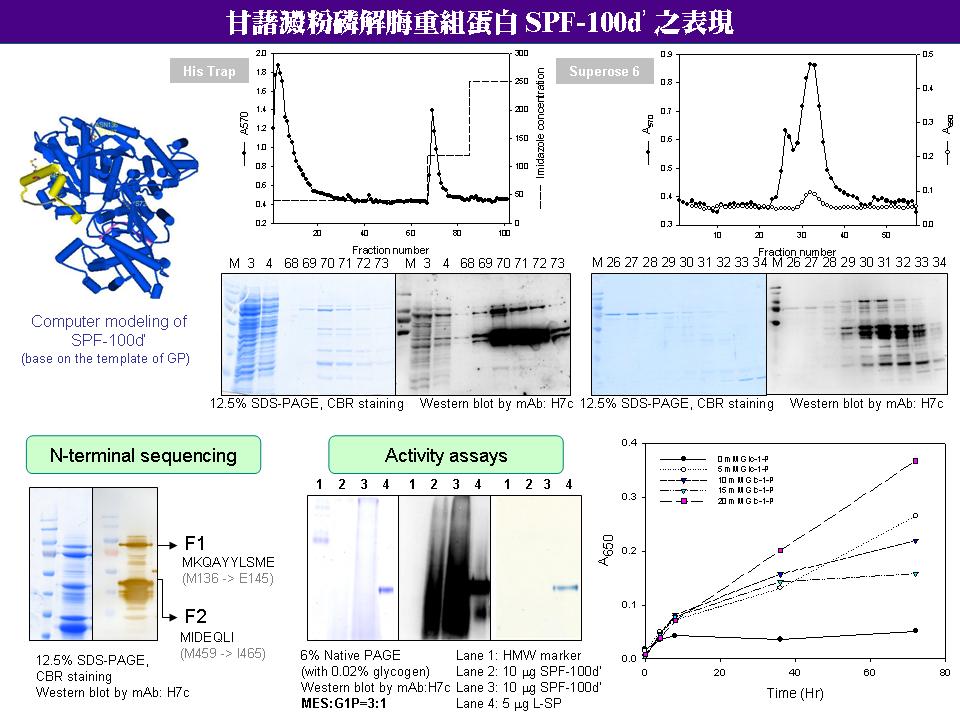
1. 用 Ni-NTA affinity column 純化出的 SPF-100d’,CBR 染色和 Western 呈色不成比例,可能是 CBR 對此表現蛋白染色較果不佳。 2. 用 E. coli 做表現蛋白,需找到表現溫度和時間的平衡點,減少蛋白質降解的比例,以使其活性增加。 3. 破菌純化蛋白質時,可以加入 protease inhibitor cocktail,減少蛋白質降解。 4. 用 gel filtration 純化 SPF-100d’ 是否每次都 2 個 peaks? 要推得表現蛋白的分子量,以了解其四級結構如何。 5. 測定表現蛋白合成方向活性時,可以用短鏈醣 (G2-G5) 取代澱粉做為 primer,再用碘液呈色法分析產物,取代磷酸根呈色。 6. L78 上有多對重覆的 glutamine 和 lysine,可能是為了抓住剛合成的短鏈醣 (G2-G4),可以計算 L78 上 EK 的分布距離,是否符合 G1-G4 的分子長度。 7. 需克服表現蛋白質活性和純化量的問題,找到穩定的活性分析條件。 8. 可測試外加 PLP 是否會對活性有幫助。 9. 可在 SPF-100d’ 酵素液中添加 L78,看是否能恢復 PI activity。 10.目前正在修改 L78 上的錯誤序列,盡速取得正確的全長進行表現。 11.以表現蛋白進行對 L-SP 活性機制的探討,除了全長和不含 L78 的部分全長之外,應針對其他功能性區城 (如 L78 上的重要 lysine) 做突變,深入分析 PI activity 可能的機制。 12.注意報告和寫作內容的打字規則,小心勿使用只有中文才能顯示的單位,然後以為它是英文 (如 ℃ 改用 ºC)。 |
| 08/08/28 |
0 金 禹 圻 豬胚胎發生的重要蛋白質表現 |
|
|
|
| summary |
1. 繼續製備動科系所提供蛋白質之抗體: 目前 o45 已接近完成,下週放大 T25 後進行腹水製備大量抗體。o48 無效價。o422 需再免疫測效價。 o432、o438、o439、o440、o42、o84、o836-1 繼續免疫及測效價。 2. 查 AK223085、BC022328、NM003333 與胚源轉換相關 paper;查 Ig k chain V-III region, arginine deiminase-like protein, lactate dehydrogenase B 與班馬魚胚源轉換的相關 paper。 3. 班馬魚 128-cell 及 512-cell 時期的 2-DE 差異。 農委會計畫剩餘款: 本月尚未報帳,全部尚餘 401,900,其中材料費餘 172,000,雜支餘 100,000,臨時工資剩 129,900。 |
| 08/08/27 |
7 林 怡 岑 L-SP 與 D-enzyme 的交互作用 |
 |
|
| summary |
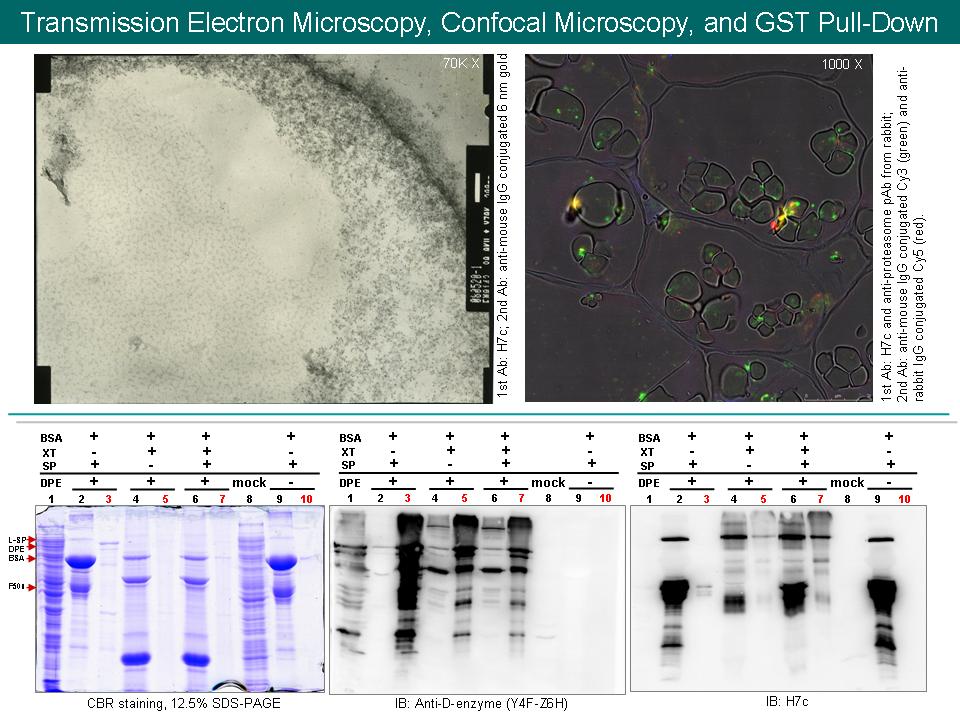
1. 儘快完成 in vitro degradation assay (+ proteasome) 與 bio-image 實驗。 2. Km 應以 mM 表示,而非以百分比表示。 3. TEM 圖有點模糊,是否能更聚焦? 4. 三張不同倍率的 confocal microscopy 組織定位,應顯示同一位置,然後再局部性放大。 5. Confocal microscopy 組織定位圖應降低 H7c (綠色) 亮度,使 L-SP 和 proteasome 重疊 (黃色) 更為明顯。 6. 為何 L-SP 和 proteasome 重疊區 (黃色) 大多位於 pellet 或細胞破片處,而且在細胞中含量很少? 7. 如何解釋 L-SP (綠色) 和 proteasome (紅色) 重疊區 (黃色) 在生理上為何會互相結合? 可能的生理作用機制? 8. 調整光學顯微鏡之光源角度,使視野更清晰。 9. 可直接在一次抗體上標定不同螢光來進行 confocal microscopy。 10. 可設計 3~5 段 D-enzyme 之 peptides 或改用接有 His-tag 的 D-enzyme 表現蛋白來免疫兔子,以防止 GST 干擾。 11. HX 經 CIAP 處理後,可利用 2D 圖譜檢測是否有去磷酸化現象。 12. 可設計實驗預測 L-SP 和 D-enzyme 的 binding site? 13. 比較在原態電泳片中,L-SP 外加 D-enzyme 所形成之高分子量色帶位置,與 HX 之高分子量色帶位置是否相同。 14. 假若 L-SP 與 D-enzyme 在 amyloplast 中互相結合,其生理角色為何? 15. 比較 L-SP only 與 L-SP 外加 D-enzyme 後,對於不同基質 Km 值之差異。 16.可在 L-SP 外加 D-enzyme 中,加入 L-SP 的基質,測定 D-enzyme 產物的生成,或是加入 D-enzyme 的基質,測定 L-SP 產物的生成,來預測可能之代謝途徑。 17. Hikaru et al. (2008) Plant Cell 20: 1833 所描述的 factor X 是否可能為 D-enzyme? D-enzyme是否會受到溫度的影響? |
| 08/08/20 |
10 何 杰 龍 禽流感病毒的免疫學工具 |
|
|
|
| summary |
1. 本研究分為三個主要方向:禽流感病毒 (AIV) 抗體庫建立、蛋白質體學研究、致病機制探討。 2. 以兩株不同亞型 AIV 病毒株為抗原,直接分離病毒顆粒進行免疫,篩出多株可以辨識 H6 亞型的專一性單株抗體。 3. 為了增加 AIV 抗體酷的多樣性,嘗試以合成 peptides 免疫,由王金和老師實驗室提供的 2838V 基因序列預測,設計一般抗原 peptides 共 24 段,高免疫性抗原 NA 共 5 段,分批進行小鼠免疫與單株抗體製備,目前持續進行。 4. 以合成 peptides 進行免疫難度很高,影響免疫結果因素眾多。為了一次排除這些無法控制的因素,將所有 29 段 peptides 混合後,同時對紐西蘭兔進行免疫。免疫後採血可辨認 3233 與 2838V 病毒 2DE 圖譜上的蛋白質。此外,我們分別將所有 peptides 吸附到不同的反應槽進行 ELISA,由此確認較易產生抗體的 peptides 片段。 5. 雖然由 competitive ELISA 結果顯示兔子血中含有辨認某些 peptides 的抗體,但一般的 peptides 合成皆含有強酸,回加以後可能會影響抗體與抗原的反應 (造成假性抑制),須再次確認 pH 值變化。 6. 因為 AIV 抗體庫的製備,除了 HA 以外其他蛋白質皆透過序列的預測與設計,因此由此研究工作,希望可以將「抗體庫」的概念轉成「免疫決定基庫」 (epitoipe bank),延伸原有的概念到 epitope 層次。 7. 致病機制研究,可由毒性不同的 2838V 與 2838N 著手,用已有 HA 的單株抗體或其他抗體,比較 2DE 圖譜差異。 8. 持續以現有抗體工具進行 AIV 蛋白質醣基化位置,對蛋白質體學及致病機制的影響。 9. 尋找 HA 上影響高低病原性與跨物種傳染的關鍵性 epitope。 10. 要注意實驗記錄本必須認真、確實整理。 |
| 08/08/13 |
20 王 信 傑 Phytochelatin synthase 的磷酸化與活性調節 |
|
slide not shown
|
|
| summary |
1. 重金屬 Cd 除了會影響 PCS 活性外,也會影響 PCS phosphorylation,本實驗發現 AtPCS1 的磷酸化是 Cd-dependent。 2. 酵素動力學實驗結果發現 AtPCS1 有 sigmoidal curve,而 AtPCS1-N 則為傳統的 Michaelis-Menten hyperbolic curve,顯示 AtPCS1 在酵素調控方面可能屬於 allosteric regulation。而且經由膠體過濾決定分子量實驗也發現 AtPCS1 為 dimer (110 kD)、AtPCS1-N 為 monomer (28kD),符合動力學結果。 3. 由 site-directed mutagenesis 實驗結果得知 AtPCS1 磷酸化位置在 Thr49,此位置突變對活性也有重大影響。另外經由分子模型預測 Arg183 位置經突變後,明顯抑制 AtPCS1 活性,推測兩者間應有重要的交互作用關係。 4. 在 AtPCS1-N 突變實驗發現當 Tyr55 突變成 Ala 後活性下降,但是若突變成 Asp 則活性上升,推論此胺基酸可能與重金屬 Cd 的結合有關。 5. 除目前已知 Cys56、His162、Asp180 為 AtPCS1 催化鐵三角外,經由分子模型預測軟體又推測出 Arg152 與 Lys185 可能與基質 (PCn) 的結合有關。 未來方向 1. 再經 site-directed mutagenesis 確認相關胺基酸的重要性。 2. 進行 truncated AtPCS1 variants crystallization 實驗,藉由結構分析來確認 PCS 催化機制。 3. 進行阿拉伯芥內生性 PCS 與其相關 kinase 或 phosphatase 純化,並探討磷酸化對生理意義之影響。 |
| 08/05/28 |
2 林 之 儀 甘藷塊根 L-SP 之 PI 活性催化機制 |
 |
|
| summary |
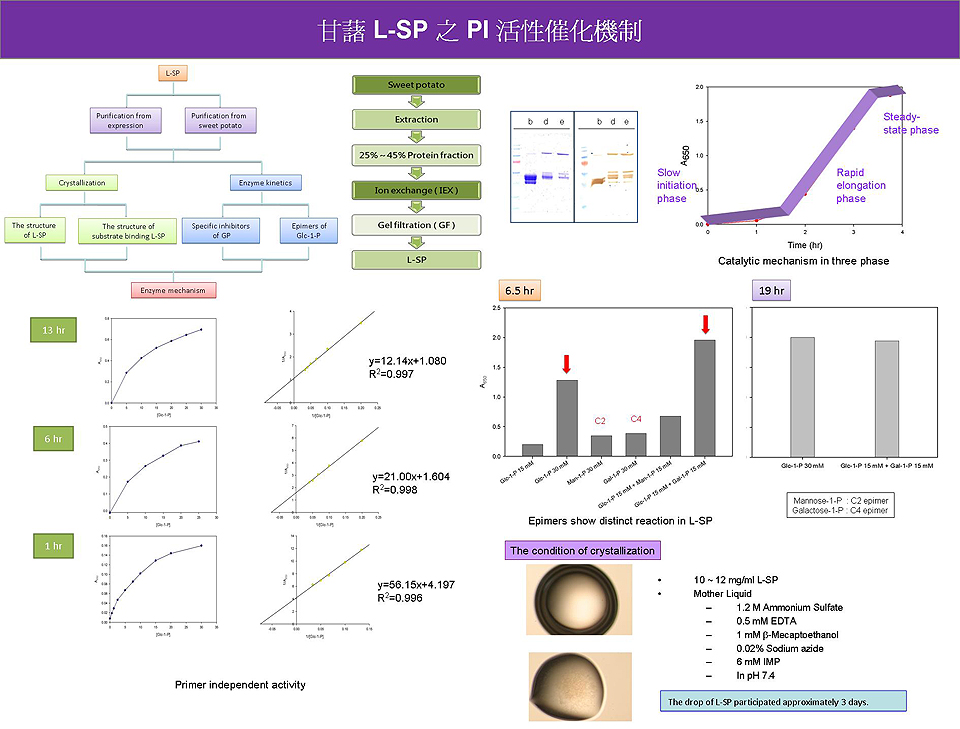
1. 重新訂定純化策略,以求快速得到大量且降解穩定的蛋白質。 2. 以相同基質量在不同時間進行 PI activity 反應,可以看到三相變化。 3. 以不同基質量在同一時間進行 PI activity 反應,則符合 Michaelis-Menten equation。 4. 進行 PI activity 的 epimer 實驗時,發現若反應時間縮短至 6 h,Glc-1-P 與 Gal-1-P 混合液的反應活性,大於只含 Glc-1-P 的基質。 5. 結晶仍在尋找最適合的 conditions。 Future work: 1. Find out the preservation conditions to control the degradation of L-SP. 2. Identifie the role of Gal-1-P in L-SP catalysis. 3. Analyze the chain length distribution during polymerization by HPLC. 4. Compare the effect to PI activity by replacing Glc-1-P to its analogs or GP inhibitors. 5. Optimize the conditions for crystallization. |
| 08/05/07 |
2 羅 庭 芳 不同禽流感病毒株之蛋白質體與毒性強弱之關係 |
 |
|
| summary |
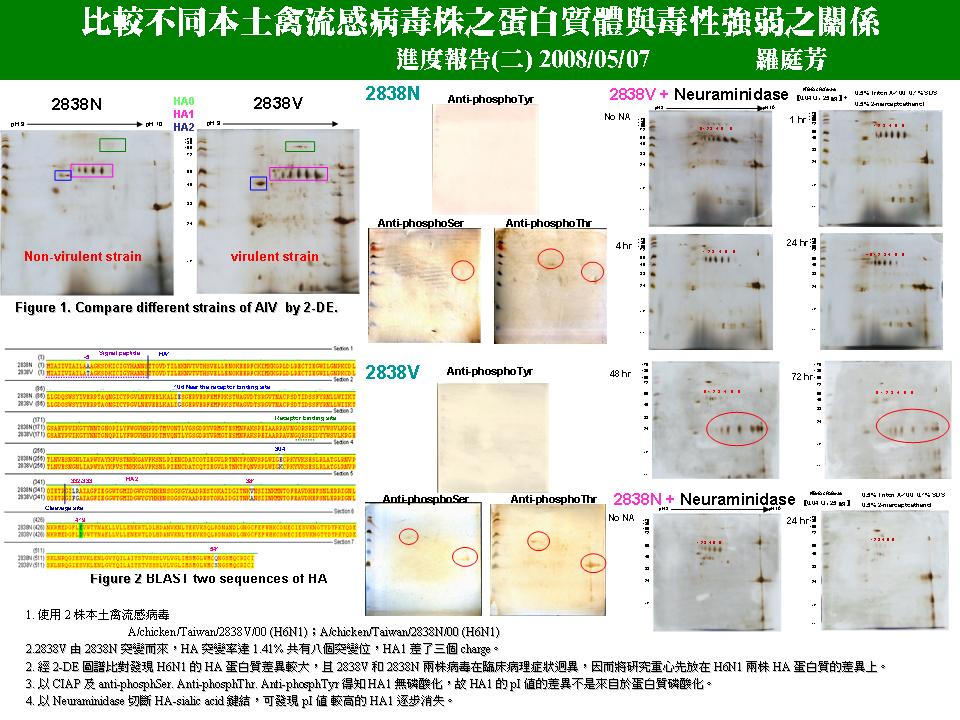
1. 以電腦分子模擬 HA 的蛋白質立體結構。 2. 再以多種方法確認 HA1 確實和 SA 鍵結。 3. 六個 HA1 不同 pI 值的色點,是否源自 HA 蛋白質與 6 個 SA 的連續結合? 4. 再整理清楚流感病毒進入宿主及離開宿主的反應細節。 5. 可否取得雞腎臟分離出的病毒?以比較由雞蛋繁殖病毒之 2DE pattern 是否有差異? (這六個點是否為雞蛋所修飾) 6. 使用 MAA 等 lectin 證明 sialic acid 的存在 (lectin binding)。 7. pI 值較高的 HA1 逐漸消失 (而低 pI 者逐漸加強),先以免疫染色確定 HA1 和 NA 作用後的位置如何。 Future work: 1. Immunoblot analysis (HA Ab) 2. Bio-mass spectrometry to identify key spots 3. Periodic acid-Schiff (PAS) identification of the AIV posttranslational modification (glycan) 4. Quantification of the HA-sialid acid binding by glycan microarray technology (N vs V) 5. Virulence of 2838N & 2838V could be distinguished by binding affinity between HA and sialic acid. 6. Cell & tissue immunostaining for target organ: renal virus |
| 08/03/26 |
4 蔡 和 成 建立差異性蛋白質體之抗體庫 |
 |
|
| summary |
1. 題目:建立綠竹筍生長過程差異性蛋白質體之抗體庫 (The Differential Proteomic Antibody Bank of the Fast Growing Bamboo Shoots)。 2. Immunoaffinity-based protein subtraction 部分要討論失敗原因: a. 抗體專一性、親和性不高 b. 兔子免疫性的問題 3. B 細胞先過 flow 或先 fusion? 跟吳建興博士討論一下實驗策略。 4. IgG & IgM 是 secretory form 或 receptor form? 兩者皆有? 5. 注意 labeling condition 及細胞離心轉速。 6. DyLight 488 是怎樣的螢光? 分子構造如何? 7. 在能夠將兩種細胞分開的狀態下,接愈少的 biotin,擋住 binding site 的機會愈低。 8. 確定綠竹筍整個蛋白質體皆能接上 biotin:比較 2D CBR 與 2D Western (用不同比例 B/P 測試)。 9. 開始後續實驗,用 total protein 開始進行 subtraction。 |
| 08/03/19 |
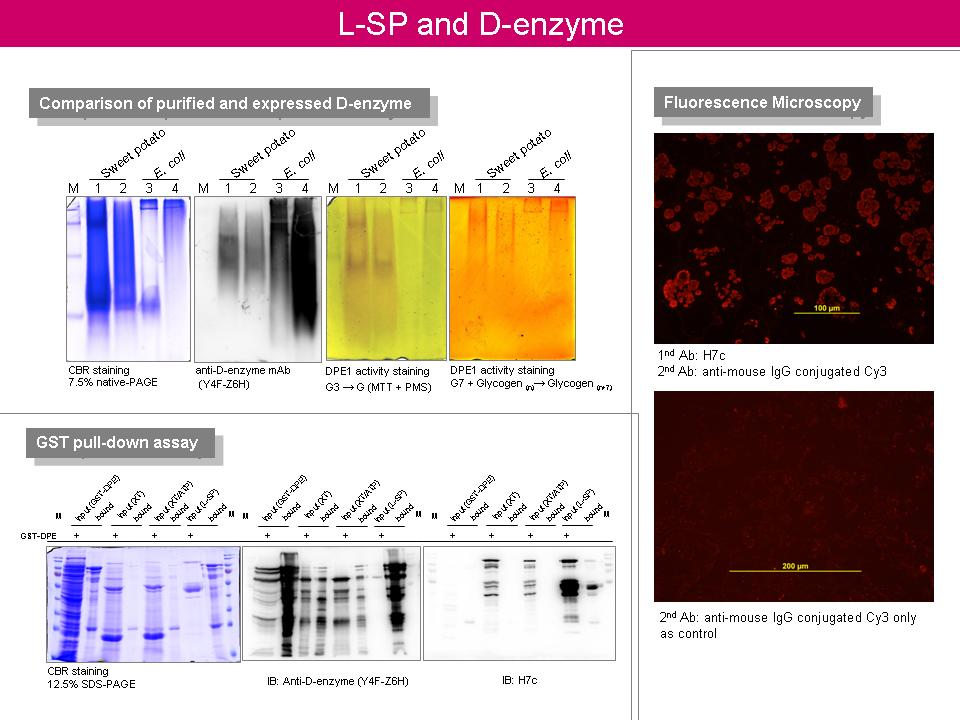
6 林 怡 岑 L-SP 與 D-enzyme 之交互作用關係 |
 |
|
| summary |
1. 免疫組織定位部分的實驗應儘速進行,可直接在一次抗體上標定 biotin 或不同螢光。 2. 瞭解 FRET 是否能應用在 confocal microscopy 上。 3. L-SP 與 D-enzyme 之基因都已取得,可考慮進行 yeast two-hybrid? 4. 為何 D-enzyme 的活性染色結果,與一般澱粉呈色不一樣? 事後查證:由於澱粉結構不同,碘液呈色也會有所差異,直鏈澱粉碘液呈色為藍黑色,支鏈澱粉碘液呈色為較淺之紅棕色。 5. 進行 GST pull-down assay 需做好 control,例如 input 之蛋白質量需適當、設計 glutathione Sepharose bead only 之對照。 6. 可否利用 GST pull-down assay 在 in vitro 下產生像 HX 這樣的高分子量活性色帶? 7. 可利用 gel-filtration 推測 HX 的分子量。 8. 可利用醣染色去觀察 HX 中,是否含有短鏈的 amylose?
9. E.
coli 中的
malP
10. malQ 之 stop codon 與 malP 之 start codon 部分重疊,那麼在分子機制上,ribosome 如何轉譯蛋白質? |
| 08/03/12 |
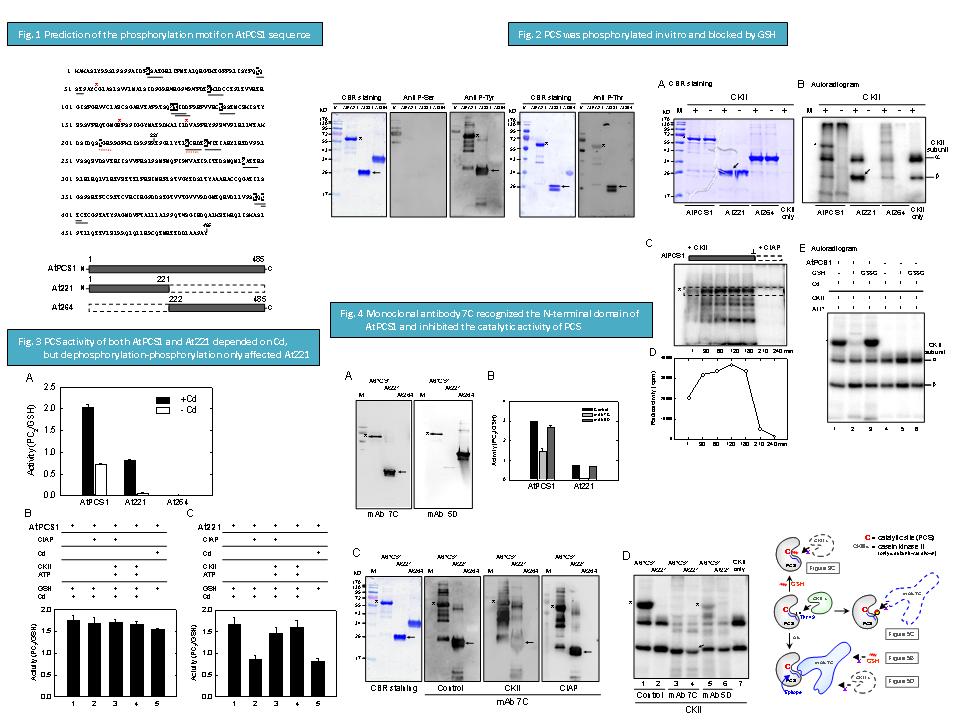
19 王 信 傑 PCS 之磷酸化與活性催化機制 |
 |
|
| summary |
1. 經由 anti P-protein mAb western blot 結果得知在 E. coli 內重組蛋白 AtPCS1 與 At221 已有部份磷酸化,且磷酸化位置可能在 Thr / Tyr 上。純化重組蛋白已有活性,後來實驗得知磷酸化促進 PCS 活性。初步推論 PCS 可以被磷酸化。 2. 進行 PCS 序列比對發現 PCS 含有兩種磷酸化保守性序列,作用酵素分別是 casein kinase 2 與 protein kinase C,再經軟體分析發現 CK2 磷酸化機率較高,以下實驗均以 CK2 磷酸化 PCS 為主。 3. 經由 CK2 放射性磷酸化實驗得知 PCS 確可被磷酸化,且磷酸化位置在 N-端 domain,另外 PCS 也可被 CIAP 作用而去磷酸化,磷酸化前後 PCS 活性也有變化 (5),顯示 PCS 可能經由磷酸化與去磷酸化來調節功能。 4. 當 GSH 與 PCS 結合後,阻斷磷酸化作用,而氧化態 GSSG 則否,推論磷酸化位置可能在 substrate binding site 附近。 5. PCS 活性需要重金屬存在才能活化,藉由 CK2 磷酸化與 CIAP 去磷酸化對活性影響實驗得知,去磷酸化會降低 PCS 活性,若再磷酸化則會回復活性,可見 PCS 確實可藉磷酸化來調控活性。 6. 由兩株專一辨認不同 PCS domains 的抗體發現,辨認 N-domain 的 mAb 7C 會抑制 PCS 活性。而 mAb 7C 與 PCS 結合後,也會阻斷磷酸化作用,推測抗體辨認位置也在活性區部份,且接近磷酸化位置。另一方面,若 PCS 先經磷酸化後,則 mAb 7C 無法辨認 PCS,更可確定抗體辨認位置可能磷酸化胺基酸上。 未來工作 1. 經由定點突變確定磷酸化位置。 2. 藉由 AtPCS1 / At221 kinetic assay 說明 PCS 可能作用機制。 3. 藉由以上實驗配合 2005 PNAS 所發表論文的 PCS 立體構造,推論出可能作用機制。 4. 進行阿拉伯芥中,內生性 PCS 之純化與分析,期能真正在植物中看到 PCS,以及其磷酸化形態。 |
| 08/02/27 |
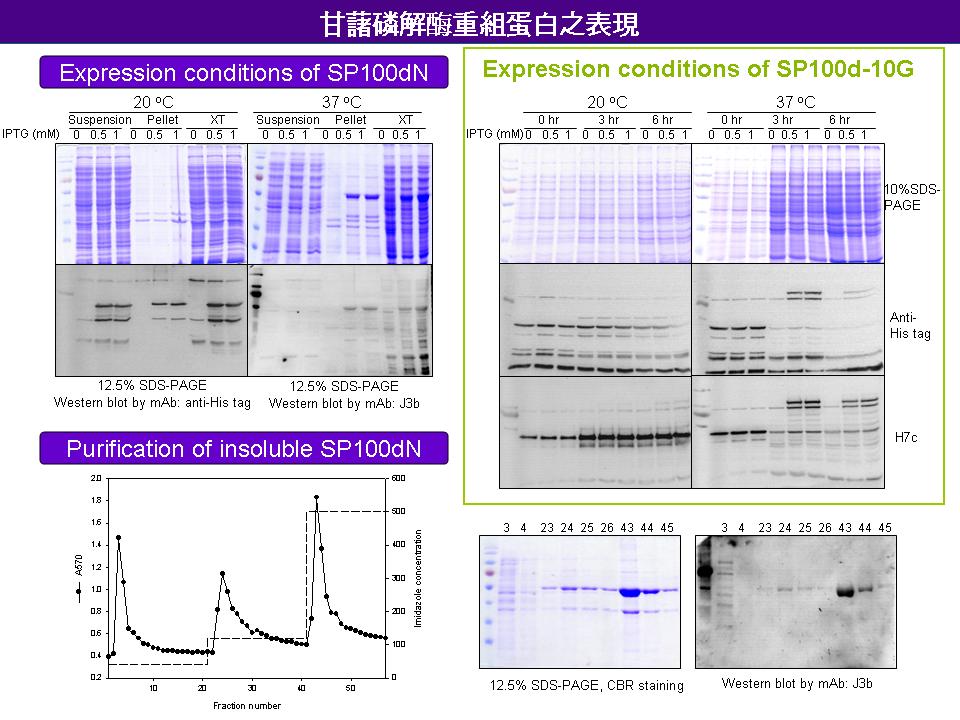
2 賈 儒 珍 甘藷 L-SP 重組蛋白之表現 |
|
|
|
|
summary |
1. 已完成構築於表現載體的片段有 SP100dN 和 SP100d-10G (L100 以 10 個 glycine 取代),保存於中間載體的片段有SPN、L100 和 SP100dC。 2. SP100dN 用 37C 誘導表現時,蛋白為不可溶性,而且 Refolding 的效果不佳;若用 20C 誘導其表現,可得到少量但可溶於水的表現蛋白。 3. J3b 辨認 SP N-端區城可能有部份靠近 L78,因此對 SP100dN 的辨認能力較差。可以用 SP100d 表現蛋白做 Western,進一步確認J 3b 的抗原辨認區是否與 L78 有關。 4. SP100d-10G 的表現量很低,正在測試其最佳表現條件。 5. SP 全長的釣取仍在進行中。由於帶有 L78 的 SPN 片段取得不易,希望由已取得的 SPN cDNA (codon 有 5 處錯誤) 修正錯誤片段,再表現 SP 全長。 6. 報告時應同時 review 原來國科會計畫內容,注意計畫進度;列出每個片段所要進行的實驗和原因,以免混淆。 未來工作 1. 測試 SP100d-10G 表現蛋白是否具有活性,若有活性將可進行 PI activity 相關實驗。 2. 構築 SP 全長和其他片段表現載體。 3. 針對功能區塊和磷酸化位置設計定點突變。 |
| 08/01/30 |
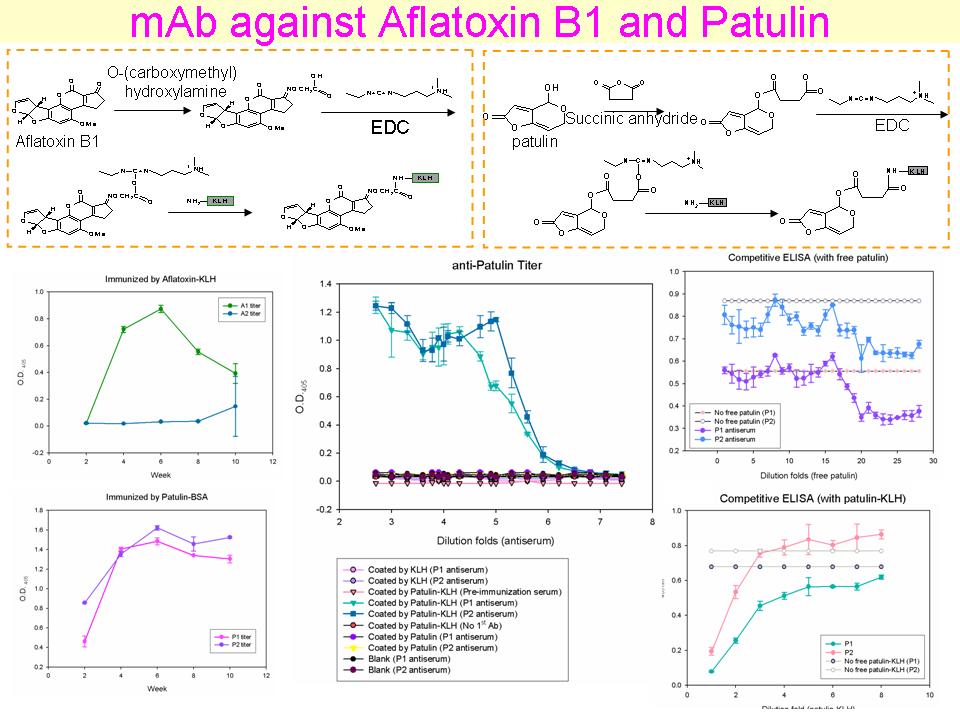
2 眭 毓 庭 小分子量毒素的專一性單株抗體 |
|
|
|
|
summary |
1. Patulin 的競爭性 ELISA 沒有抑制效果,但是 KLH-patulin 卻可看出抑制效果。 2. 可能 patulin 樣本壞掉,解決方法: a) Patulin 要分裝,避免重複解凍、b) Patulin 樣本中可能有其他干擾物質、c) 用 H2O 取代 PBS。 3. 也可能抗體結合的不是 patulin 的 native form,而認到 linker 或是 modified patulin (patulin-linker)。 4. 可能的解決方法: a) 設計其他 conjugate 方法,讓 linker 短一點,或用 affinity gel (Sepharose) 接 patulin 或將 patulin 聚合。 b) Patulin 和一抗直接加入 coating 好的 plate 中反應。(這好像沒有什麼用) c) 先用 succinimide anhydride 和 patulin 反應,作為 antigen 與一抗結合,看是否抗體認的是 modified patulin。 d) 在抗體與 patulin 反應時,將抗體和 patulin link 固定起來 (抗體 knockout),確保抑制性的反應。 5. 在效價測試方面,血清稀釋 25,600,000 倍不合理,稀釋倍數過高,血清中應該已經沒有蛋白質。 |
| 08/01/23 |
3 劉 雨 亭 主題 |
|
|
|
|
summary |
|
| 08/01/16 |
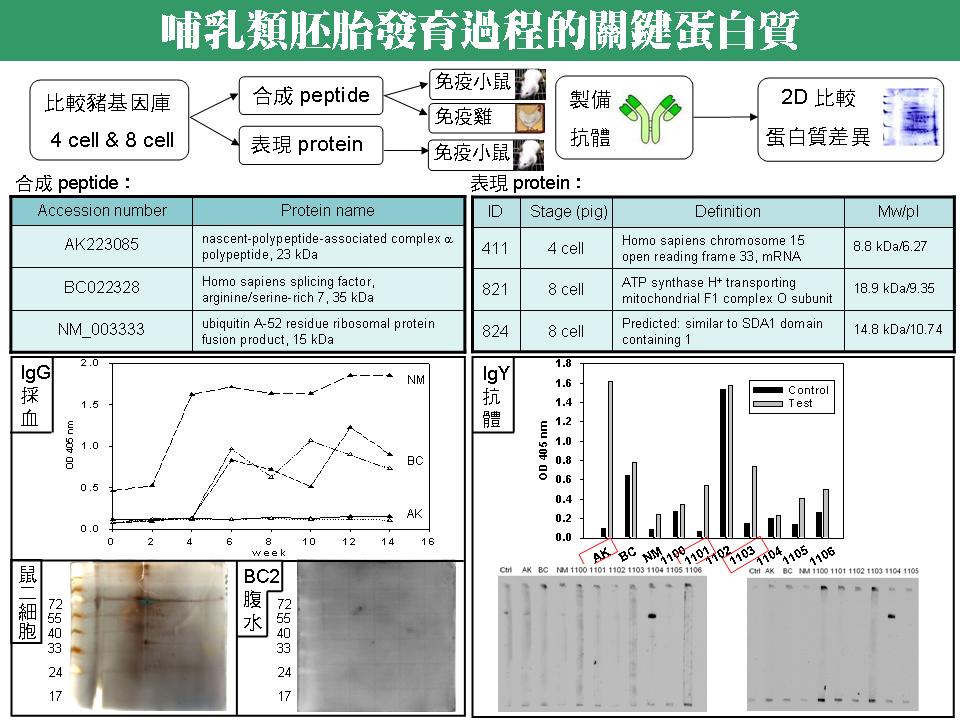
3 沈 志 昱 建立哺乳類胚胎發育過程中關鍵性蛋白質之抗體庫 |
|
|
|
|
summary |
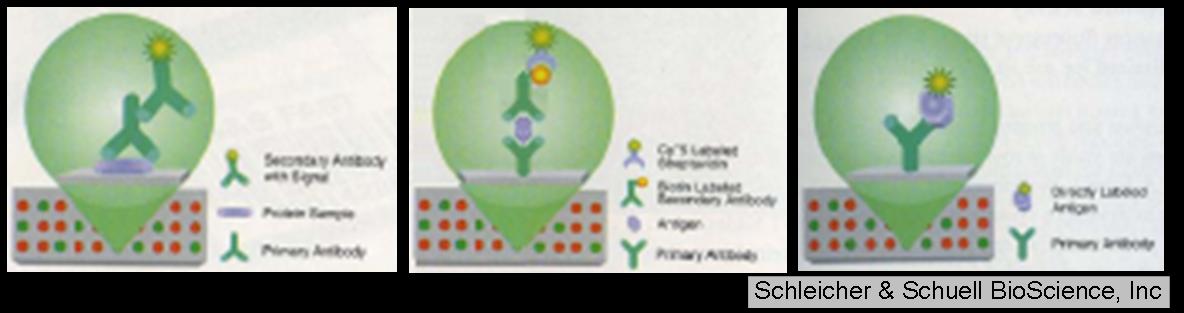
1.現有抗體之整理: IgG 方面:採血 (AK1, AK2, BC1, BC2, NM1);腹水 (BC2, 824, 821, 411) IgY 方面:AK, BC, NM, 1097, 1098, 1099, 1100, 1101, 1102, 1103, 1104, 1105, 1106 預測有效價者:ELASA 測試有效價者 BC1, BC2, NM1 (採血 IgG); BC2 腹水, 824, 821 (IgG 腹水); AK, 1101, 1103 (IgY) 經小鼠 2-cell 期胚胎測試有效價: BC2 腹水, 1104 由於胚胎累積的問題,以上許多抗體尚未回歸到小鼠胚胎 Western 測試。 2. 由小鼠 2-cell 期胚胎之 2-DE Western 測試可看出,BC2 腹水似有明顯效價,且其中一色點的分子量,與預測蛋白質分子量接近 (35 kDa),然而這實驗僅做過一次,需多次確認。若比較小鼠 4-cell 期胚胎 2-DE Western,發現某蛋白質確實有消長現象,則要趕緊確認其身分,若與預測蛋白質相同,相信此抗體將有重要應用性。 3. 比較 IgY 的 ELISA 效價,及小鼠 2-cell 期胚胎測試,可發現在 ELISA 測試上 AK, 1101, 1103 三種抗體,雖具有明顯的效價,然而在小鼠 2-cell 期胚胎幾乎沒有效價。此外,1104 抗體在胚胎測試有很強的效價,但是在 ELISA 測試卻沒有明顯效價,這是比較令人擔心的地方。還是必須把 2-cell 期與 4-cell 期胚胎 Western 圖譜都做出來才能確定。 4. 小鼠胚胎實驗結果,最終仍須回歸到以豬胚胎測試,然而豬胚胎的取得非常不易,數量也少很多,無法以蛋白質體角度切入,因此得想另外的切入辦法。 三種解決豬胚數量過少的辦法:Microarray Western, Sandwich assay, Direct label 依序如下圖:
a. Microarray Western 是將特定時期豬胚胎固定在晶片表面,依序通過由小鼠或蛋雞製備的一次抗體,及帶有標定之二次抗體 (HRP 或 Cy3/Cy5),再偵測訊號。 b. Sandwich assay 是先在晶片表面接上由小鼠或蛋雞製備的抗體,將特定時期豬胚胎通過晶片表面,再通過來自兔子製備的抗體及其帶有標定之二次抗體 (HRP 或 Cy3/Cy5),同樣偵測訊號。 c. Direct label 是在晶片表面接上由小鼠或蛋雞製備的抗體,再通過直接標定 Cy3 的 4-cell 及 Cy5 的 8-cell 豬胚胎蛋白質,直接偵測訊號。 未來工作: 1. 將製備出的各種抗體,回到 2-DE 圖譜上偵測,並將各色點挖出進行 mass 定身分。 2. 持續收集不同時期鼠胚,以二次元電泳檢定後,將不同消長之差異色點鑑定身分。 3. 做出小鼠 2-cell 與 4-cell 的 2-DE 免疫圖譜。 4. 搜尋與繁殖力相關的蛋白質,這次不從資料庫挑選,直接從已發表文章找尋影響早期胚胎發育的關鍵蛋白質。 5. 試著用上述三種晶片方式去分析豬胚胎,根本解決豬胚胎樣本不足之問題。 6. 建立哺乳動物早期胚發育之抗體庫。 |
| 08/01/09 |
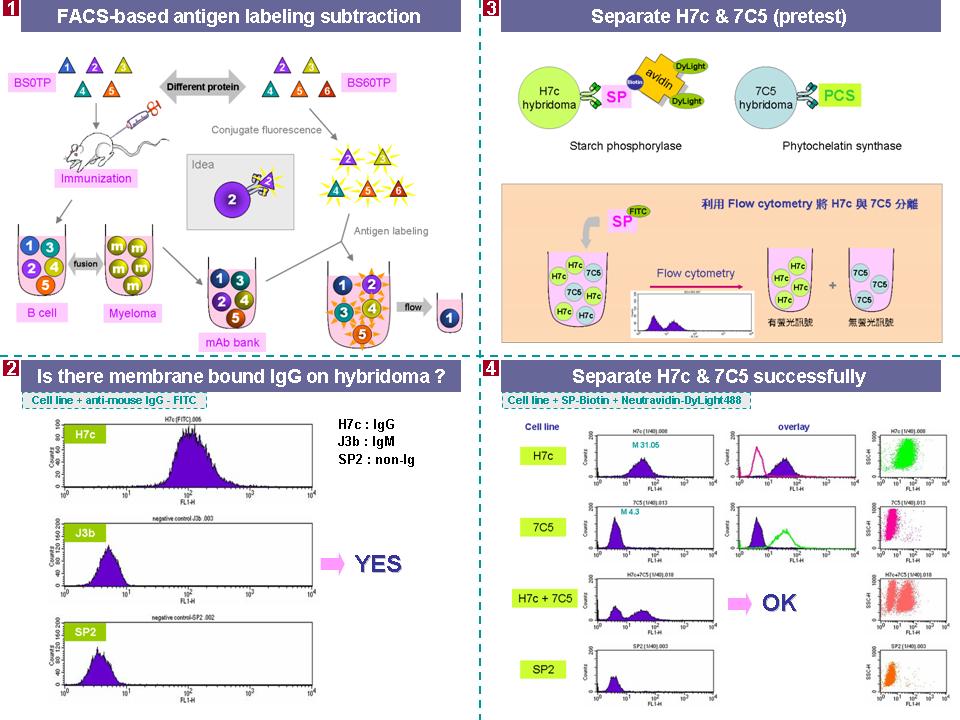
3 蔡 和 成 以流式細胞儀建立差異性抗體庫 |
|
|
|
| summary |
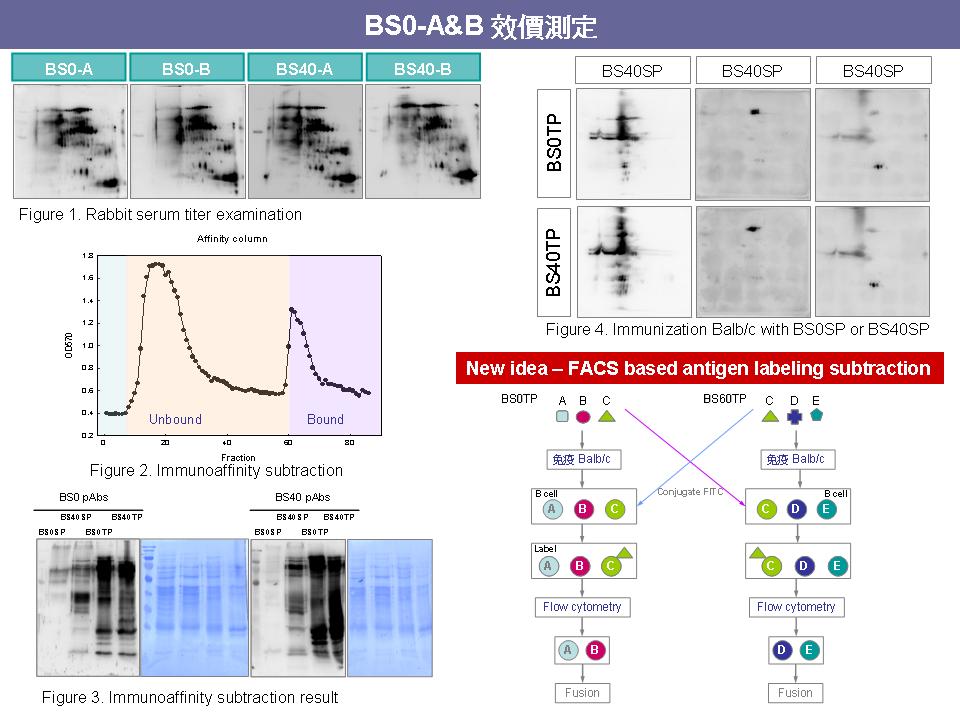
Method 1 Subtractive immunoaffinity column 1. ELISA antibody coating 時,所用的 buffer pH 改成 8~9,coating 效率較高,要做 negative 及 positive controls。 2. Sandwich ELISA 中的 rabbit pAb 是對抗全部蛋白質,無法特別在意某一抗原的 titer 高低。 3. 過完 immunoaffinity column 後,應該計算 loading 量及最後所得到 unbound 及 bound 數量。 4. R1 及 R1L1 雖然免疫相同蛋白質,但產生的免疫反應有相當差異。 5. 最後仍然繼續進度 fusion,進行抗體篩選,儘量去撈一些抗體 (順便練習)。 Method 2 FACS-based antigen labeling subtraction 1. FITC (FW 389) 標定是否會擋住 binding site? 是否可用較小的螢光物質(Cy3 or Cy5) 去標定? Ans:標定 FITC 時可以控制標定的比例,不要在一個蛋白質上接上太多 FITC,擋住 binding site 的機會就會降低。而 Cy3 無法用 488 nm 波長激發,所以無法使用。 2. 過完 FACS 的 B cell 會很少,這樣可能造成 fusion ratio 不好,且 B cell 放太久可能會死亡。 3. Fusion 後再過 FACS 也可以試 (fusion 完後先養在 T80,待細胞穩定再過 FACS)。 4. T cell 和 B cell 無法分開,全部一起進行 fusion。 5. 過 FACS 後的細胞也含有產生其他抗體的細胞 (原先因病誘出的 B cell),仍然要進行抗體篩選,但和傳統抗體庫相比,工作量降低。 6. 處理抗原及標定時盡量無菌,最後可用 filter 過濾。 |
| 08/00/00 |
1 報 告 者 主題 |
|
slide |
|
| summary |
|
{kind=link}