|
|
酵素化學實驗 B 參考講義 | ||
|
|
B4 相關研究計畫 |
|
|
| 流程圖 | |||
| 摘 要 研究計畫之背景及目的 (研究背景 研究動機 參考文獻) | |||
|
國科會研究計畫 |
|||
|
甘藷澱粉磷解脢激脢之純化與性質檢定 |
|||
|
摘 要 |
|||
|
|
澱粉磷解脢 (Starch phosphorylase, SP) 一般認為是植物分解其貯藏性多醣的重要酵素,其蛋白質是由兩個 110 kDa 次體所組成的二元體,原態分子量為 220 kDa。 與動物的同源酵素 肝糖磷解脢 (glycogen phosphorylase, GP) 比較,SP 分子的中央多出一段由 78 個胺基酸組成的 a 螺旋 (L78);L78 可能是由 intron 演變而來,剛好插入 SP 與基質結合區的正中位置,阻礙了澱粉與酵素之結合,可能因此而降低活性。 在研究 SP 的過程中,發現 SP 分子非常容易斷裂,成為一群分子量約為 50 kDa 的片段 (F50);推測可能是在 L78 上的 PEST site 上斷開。但 SP 分子在斷開後,仍保有其完整的四級構造,且其催化活性與基質的親和力都有增加。 已知動物細胞內的 GP 有很複雜的酵素調節作用,如磷酸化、異位脢調節等,但 SP 分子構造上則缺乏這些特徵;我們推測 L78 的存在,及其易受水解的特性,很可能是植物細胞用來調節 SP 活性的一種機制。 以電腦程式 PC/GENE 搜尋 L78 上的胺基酸序列,發現有很強的磷酸化位址,經單株抗體檢定確有磷酸化 Ser,而以放射性 ATP 標示,也發現 SP 分子可以被接上放射性磷;另外在 L78 的 C-端側發現有 Pro-rich 序列,可能與 SH3 domain 有專一性結合,而 SH3 是信息傳導的重要聯結分子。我們也發現 SP 可以在沒有引子的存在下,使用 Glc-1-P 合成長達數千葡萄糖單位的直鏈澱粉。因此,我們擬由 SP 的磷酸化,以及 SP 可能參加的信息傳導網路,推知植物細胞中澱粉顆粒生合成的誘導,與其細胞層次的調節控制是如何發生。 由於我們已確知SP會受到磷酸化,因此甘藷塊根中一定有可以進行磷酸化的激脢存在。因此,本年度計畫將要接著完成下面三項工作: (1) 純化此 SP 激脢並檢定其生物化學的基本性質; (2) SP 的磷酸化是否確實與信息傳導有關; (3) 製備 SP 激脢的抗體以作為檢定的方便工具。 |
||
|
研究計畫之背景及目的 |
|||
|
|
|
一、本計畫之研究背景: |
|
|
動物的肝糖磷解脢有極為複雜的調控機制: |
|||
|
以貯藏性多醣類為基質的磷解脢 (phosphorylase),是生物體內重要的醣類代謝酵素 (Fletterick and Madsen, 1980)。 尤其在動物細胞中,肝糖磷解脢 (glycogen phosphorylase, GP) 影響動物體內血醣濃度的高低,有極複雜的酵素調節及控制系統,數十年來被研究得很透澈,有關的重要結論例如: |
|||
|
a. GP可經磷酸化,由不具活性的 b 型轉變為活性 a 型;反之則由去磷酸反應,變為不具活性的 b 型。 磷酸化的位置在 Ser 14。 |
|||
|
b. GP 有許多 allosteric effectors,如 caffeine 及葡萄糖均為抑制性影響;而 AMP 則為 activator。 都可誘導蛋白質分子構形的改變,造成抑制或活化效果。 |
|||
|
c. 肝糖磷解脢對基質有一結合位置,距活性區 30 埃,方便與澱粉結合,GP 的細部分子構造已經由 X 光繞射分析解出 (Acharya et al., 1991; Johnson and Barford., 1990)。 |
|||
|
植物的澱粉磷解脢很容易降解但仍保持其原態四級構造: |
|||
|
對等於動物的 GP,澱粉磷解脢 (starch phosphorylase, SP) 則被認為是植物分解其貯藏性多醣的重要酵素 (Steup, 1988),本研究群體已從甘藷塊根中純化此酵素 (Chang et al., 1987),群殖得到其 cDNA 及 genomic DNA 並且定序完成 (Lin et al., 1991)。一般的生化研究,則檢定 SP 的蛋白質四級構造,是由兩個 110 kDa 次體組成的二元體,故原態分子量為 220 kDa (Chang et al., 1987)。 在研究 SP 的過程,發現 SP 分子非常容易降解,可在 SDS-PAGE 上得到一群分子量約 50 kDa 的片段 (F50),推測可能是 SP 在分子的中央裂解所得 (Chiang et al., 1991);而 SP 分子雖然斷裂,但仍然可維持其三、四級構造,而具有原來的催化活性。 我們也對 SP 製備得單株抗體多株,發現不同的單株抗體會與不同的 SP 片段結合,在判定分子的斷裂點及其他研究工作上極為有用 (Chern et al., 1990)。 Brisson 等也檢定馬鈴薯 SP 的基因表現,發現其 SP 可能也受到轉譯後之調節 (St-Pierre et al., 1996)。 |
|||
|
澱粉磷解脢分子中央多出一段由 78 個胺基酸組成的多汰: |
|||
|
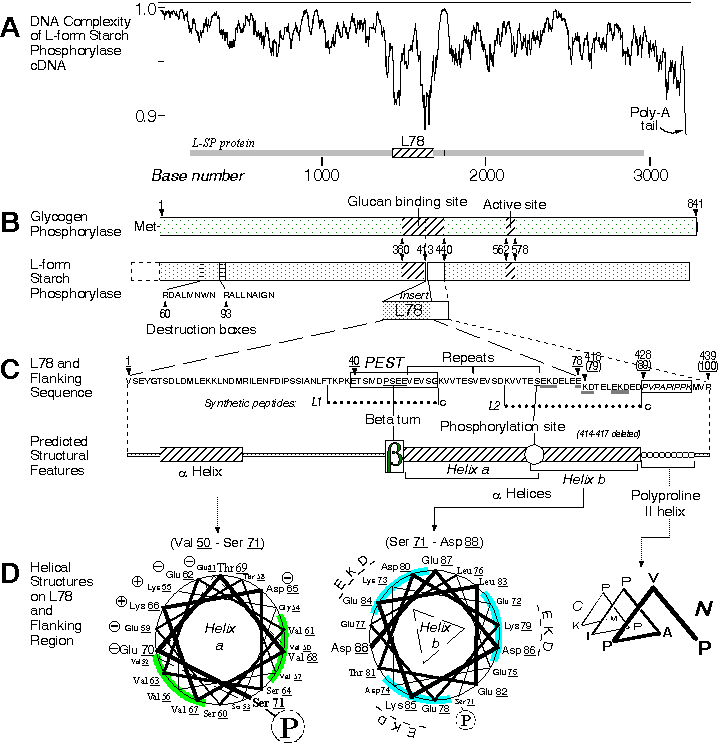
日本大阪大學的福井等人 (Nakano and Fukui, 1986),群殖了馬鈴薯的 SP cDNA,並由其胺基酸一級構造預測 SP 分子的三次元結構。 與 GP 比較後發現,SP 與 GP 的立體構造相當相似,但在分子的正中央,多出一個 78 胺基酸的 a 螺旋環 (L78)。 此環位置剛好切入上述 GP 的多醣結合區,且突出分子表面,造成了 SP 對澱粉結合的障礙。 此外 GP 所有的磷酸化、allosteric effect 等作用,在 SP 上均不存在;因此 SP 不像 GP,沒有明顯的調節機制。 以 PC/GENE 分析 SP 的胺基酸序列,我們發現在 L78 分子的正中央,有一很強的 PEST site;表示其半衰期很短,可能肇因於蛋白脢對 L78 的攻擊。 以上有關的分子構造關係,整理於 圖一。 福井也由馬鈴薯中群殖得 SP 的 H 型異構脢 (Mori et al., 1991),H 型 SP 分子中因無 L78,故對肝醣的和力較高;他們把 L 型中的 L78 片段植入 H 型中,發現修改後的 H 型 SP 對澱粉的親和力因而下降 (Mori et al., 1993)。本群體亦發現 b-澱粉脢 會抑制 SP 的活性,可能與基質的競爭有關 (Pan et al., 1988)。 |
|||