你知道台大有「甲殼類病毒學研究室」嗎?
動物系 郭 光 雄.羅 竹 芳
| No. 3 (September, 2000) | ||
|
|
||
|
生物醫學報導 3 | |
| 台大甲殼類病毒學研究室 | ||
|
你知道台大有「甲殼類病毒學研究室」嗎? |
||
|
動物系 郭 光 雄.羅 竹 芳 |
||
|
|
||
|
鮮有人知道本校動物學系有一專門從事甲殼類病毒學研究的實驗室,但若提到魚類病理學、昆虫桿狀病毒表現載體系統及蝦類白點症病毒等研究,可能就會有不少的學者專家或養殖業者聯想到台大動物系我們所共有的研究室,也就本文所要介紹的甲殼類病毒學研究室。 |
||
|
|
||
|
圖一 罹患病毒性白點症之熊蝦 |
||
|
|
|
|
|
草蝦、紅蟳、大閘蟹等盤中佳餚均屬於甲殼類動物,而甲殼類病毒學研究室就是專門研究感染這些蝦蟹類的病毒,其中尤致力於蝦類白點症病毒 (white spot syndrome virus, WSSV) 之研究。早自 1993 年起,中國大陸之養殖蝦類開始流行一種會造成蝦群大量死亡的病毒性疾病,由於罹病蝦體之外骨骼 (蝦殼) 上經常出現白點 (圖一),因此被稱之為白點症 (Wang et al., 1995)。隨後白點症以極快的速度蔓延至全球,使世界養蝦產業的成長頓時停滯。毫無例外地,本省養蝦產業亦受到嚴重波及,致使養蝦業者一提及白點症便有「談虎變色」之感。根據估算,近年來亞洲地區養蝦產業因白點症之流行,每年的損失約達到 10 億美元,由於蝦類向來為台灣重要的水產養殖項目,而白點症的出現無疑地造成極大威脅,鑑於此當前危機,本研究室於 1994 年底將昆蟲桿狀病毒分子生物學及鰻魚微孢子蟲致病機制等研究路線關閉,把所有的人力及設備投入蝦類白點症病毒的研究,同時亦將實驗室更名為甲殼類病毒學研究室。本實驗室所進行之研究層面涵蓋了分子 (Tsai et al., 2000a; 2000b; van Hulten et al., 2000)、細胞 (Wang et al., 2000)、病毒 (Wang et al., 1995; Lo et al., 1999)、病毒宿主 (Lo et al., 1996b; Chen et al., 2000)、養殖環境、天然種蝦資源 (Lo et al., 1997)、蝦類繁養殖技術及疾病控制的理論 (Lo et al., 1996a) 與應用 (Tsai et al., 1999; Hsu et al., 2000) 等,以期全面了解白點症病毒的感染及致病機制,並早日建立蝦類白點症之控制策略。 |
|
|
|
蝦類傳染性的疾病包括病毒、細菌、立克次氏體及寄生蟲之感染症,其中以病毒感染症對養蝦產業的危害最大,特別是病毒性白點症。何謂蝦類之病毒性白點症 (virus associated white spot syndrome)? 罹患此症蝦體之外骨骼 (蝦殼) 上經常會出現白點或白斑 (圖二),其中又以頭胸部最為明顯,發病期為 2-10 天,死亡率可高達 100%,傳播迅速,是一種相當可怕的病毒性傳染疾病。目前尚無任何報導指出有任何一種蝦類對此症具有抵抗力 (Lo et al., 1996)。 |
||
|
|
||
|
圖二 罹患病毒性白點症之熊蝦頭胸甲 |
||
|
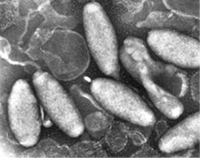
由原位雜合法 (in situ hybridization) 的分析結果顯示,白點症病毒是一種會侵襲宿主多種組織器官的系統性病原體 (Lo et al., 1997),主要侵害之組織器官包括表皮、皮下組織、造血組織、類淋巴器官、鰓、食道及胃的表皮及結締組織、中腸 (圖三 )、神經、綠腺、心臟、肌肉、複眼、及生殖細胞 (圖四) 等。被感染細胞之細胞核都呈現肥大的現象,且核內有嗜鹼性包涵體 (basophilic inclusion) 。病變細胞中出現大量具封套 (enveloped) 但不會形成封埋體之桿狀病毒 (non-occuluded rod shaped virus) (圖五)。純化的病毒為橢圓形、紡綞形或桿形 (圖六)。經負染處理之病毒粒子寬度為 70-150 nm,長度為 250-380 nm。部分病毒粒子的一端可見到尾狀突起的構造。不具病毒封套之核蛋白鞘 (nucleocapsid) 直徑為 58-67 nm,長度為 330-350 nm。在其表面可觀察到蛋白鞘組成次單位所形成之特殊橫紋構造,寬度為 20 nm,並與病毒粒子的縱軸成 90。角,是為白點症病毒之特徵 (圖七)。 |
||
|
|
||
|
圖三 呈現原位雜合正反應之感染病毒的細胞 圖四 被病毒感染的蝦卵細胞 |
||
|
|
||
|
圖五 被感染細胞中的病毒 圖六 純化的完整病毒粒子 圖七 病毒核蛋白質鞘 |
||
|
鑒於本病毒對產業之衝擊過於重大,應優先設法加以控制,雖然有關本病毒之研究報導不少,但大都止於病情、病毒電顯觀察及組織病理觀察等,對於解決產業問題之助益有限,然此時若能建立一套具有高靈敏性、高專一性、操作簡易之快速檢測診斷系統,不僅能準確及快速地篩檢出外觀正常之帶原蝦苗及池蝦,以有效防止白點症病毒的入侵及傳播,同時在配合一系列的病毒基礎生物學研究下,說不定能於可預期之時間內建立控制本病毒流行及防止其肆虐的策略。所以本研究除從事白點症病毒之致病機制及病原體分子生物學的研究外,同時亦開發核酸診斷試劑,建立高專一性白點症病毒 PCR 檢測系統 (Lo et al., 1996; 1999)。本檢測系統,已經由國科會轉移至民間企業公司生產及商品化,目前行銷國內外等 13 個國家。除廣受國內業者喜用外,世界上養蝦重鎮如泰馬及中南美洲各國養殖業者,皆廣用本系統以控制白點病的發生。 |
||
|
依 1999 年資料,顯示泰國在使用本檢測系統以控制白點病的發生後,已將每年因白點症所造成草蝦養殖的損失控制在 5% 以下,與過去之 20-70% 相比較,已大幅降低。另外,本研究利用此 PCR 檢測系統,對病毒之宿主域、組織向性分析及傳染途徑等進行探討,發現本病毒之宿主域相當廣,例如池中的橈腳類、水中昆蟲及十足目生物中的甲殼類 (蝦蟹等) 皆為白點症病毒之宿主 (Lo et al., 1998)。同時對於沿海種蝦的帶原檢測顯示,種蝦的帶原率最初似與季節之變化而有所不同,然這一兩年來,似乎已無季節性之差異,平均之帶原率為 80%。由於此病毒具有水平及垂直之傳染能力 (Tsai et al., 1999),加上草蝦養殖仍完全仰賴天然捕獲種蝦以生產蝦苗,所以在無法取得大量未受病毒感染的種蝦下,幾乎無法生產無病毒之蝦苗,同時沿海水域及養殖池一旦被病毒污染,就難以清除。此外白點症具有快速發病的特性,輕微帶原之池蝦在緊迫環境下,如水溫、鹽度之驟變、缺氧、密度過高等,都有可能在一夕之間引爆本病症,致使池蝦全數死亡。雖在如此重重困難的環境條件下,本研究室仍由所得之研究成果確立了對抗本病毒之養蝦基本條件:(1) 利用本研究室開發並透過國科會技術轉移之白點症病毒檢測系統檢測種蝦,生產蝦苗及監視池蝦帶原之狀況;(2) 以放養無白點症病毒之蝦苗為原則,若放養輕度帶原之蝦苗,則此蝦苗之帶原率不得超過 30%;(3) 蝦苗之放養密度不得超過每公頃 50 萬尾;(4) 嚴格之養殖管理,例如:避免帶原生物的進入、避免使用未經檢測之生物餌料或生物製劑、注意水溫、鹽度、及溶氧量之變化及餵飼良質之飼料等。 |
||
|
由於本研究室積極解析本病毒之基因體,目前已找出本病毒之 rr, tk, tmk, pk 及 tf 等與病原性相關的基因,現正積極分析這些基因的功能,進而探討其對宿主間之關係,除了期望在最短之時間內能找出防治本病毒症之藥物外,甚至能產生抗本病毒之基因轉殖蝦。本研究室也為著落實生產基因轉殖蝦,而積極與研發池中培育種蝦及有效的人工催熟方法。 |
||
|
隨著病毒學及分子生物學研究的快速進展,科學家們已注意到海洋生物病毒基因的獨特性,目前已有許多新的基因被發現,例如在綠藻病毒中就發現了新的限制酵素等等,由於這些基因的發現不僅對於生物體基因表現及調控機制的研究有所助益,同時亦蘊藏極大的應用發展潛力。根據本實驗室對於蝦類白點症病毒基因的研究結果顯示,此病毒之基因結構相當獨特,與已知病毒的相似性極低,屬於一種新的甲殼類病毒,因此當我們致力於白點症病之毒基因結構及功能分析時,不僅能對於此病毒之基因表現特性及其分類地位等研究有所貢獻,同時應會發現許多極具研究價值及應用潛力之基因,或許可為台灣在國際上甲殼類病毒學研究領域及應用產業方面開創新的契機。 |
||
|
就白點症病毒的基因體大小而言,估計其應具有百種以上的基因,非本研究室可以獨立完成所有基因的分析工作,所以有必要與其他研究室合作以快速進行各類基因及其產物的分析,例如蛋白質結構、蛋白質功能、酵素特性解析等等。目前本研究室己和中央研究院生化研究所黃銓珍博士、台灣大學醫學院生化研究所林榮耀院士及張智芬教授建立長期合作關係,期望藉由大家共同努力,以系統性的分析法,在最短時間內了解蝦類白點症病毒基因的結構與功能,並鑑定出有開發潛力的新穎酵素,提升台灣在生物產業上之國際競爭力,在基因資訊爆炸的時代占有一席之地。 |
||
| 參考文獻 | ||
|
1. Tsai, M.F., Yu, H.T., Tzeng, H.F., Leu, J.H., Chou, C.M., Hung, C.J., Wang, C.H., Lin, J.Y., Valk, J.M., Kou, G.H. & Lo, C.F. (2000) Identification and Characterization of a shrimp white spot syndrome virus (WSSV) gene that encodes a novel chimeric polypeptide of cellular-type thymidine kinase and thymidylate kinase. Virology (In press) 2. Tsai, M.F., Lo, C.F., van Hulten M.C.W., Tzeng, H.F., Chou, C.M., Huang, V.J., Wang, C.H., Lin, J.Y., Valk, J.M. & Kou, G.H. (2000) Transcriptional analysis of the ribonucleotide reductase genes of shrimp white spot syndrome virus. Virology (In press) 3. van Hulten, M. C. W., Tasi, M.F., Schipper, C.A., Lo, C.F., Kou, G.H. & Vlak, J.M. (2000) Analysis of a genomic segment of white spot syndrome virus of shrimp containing ribonucleotide reductase genes and repeat regions. J. Gen. Virol. 81: 307-316. 4. Chen, L.L., Lo, C.F., Chiu, Y.L., Chang, C.F. & Kou, G.H. (2000) Natural and experimental infection of white spot syndrome virus (WSSV) in benthic larvae of mud crab Scylla serrata. Dis. Aquat. Org. 40: 157-161. 5. Hsu, H.C., Lo, C.F., Liu, K.F., Su. M.S. & Kou, G.H. (2000) Studies on effective PCR screening strategies for white spot syndrome virus (WSSV) detection in Penaeus monodon brooders. Dis. Aquat. Org. 39: 13-19. 6. Wang, C.H., Yang, H.N., Tang, C.Y., Lu, C.H., Kou, G.H. & Lo, C.F. (2000) Ultrastructure of white spot syndrome virus development in primary lymphoid organ cell cultures. Dis. Aquat. Org. 41: 91-104. 7. Lo, C.F., Hsu, H.C., Tsai, M.F., Ho, C.H., Peng, S.E., Kou, G.H. & Lightner, D.V. (1999) Specific genomic DNA fragment analysis of different geographical clinical samples of shrimp white spot syndrome virus. Dis. Aquat. Org. 35: 175-185. 8. Tsai, M.F., Kou, G.H., Liu, H.C., Liu, K.F., Chang, C.F., Peng, S.E., Hsu, H.C., Wang, C.H. & Lo, C.F. (1999) Long-term presence of white spot syndrome virus (WSSV) in a cultivated shrimp population without disease outbreaks. Dis. Aquat. Org. 38: 107-114. 9. Lo, C.F. & Kou, G.H. (1998) Virus associated white spot syndrome of shrimp in Taiwan : A review. Fish Pathology 33: 365-371. 10. Lo, C.F., Ho, C.H., Chen, C.H., Liu, K.F., Chiu, Y.L., Yeh, P.Y., Peng, S.E., Hsu, H.C., Liu, H.C., Chang, C.F., Su, M.S., Wang, C.H. & Kou, G.H. (1997) Detection and tissue tropism of white spot syndrome baculovirus (WSBV) in captured brooders of Penaeus monodon with a special emphasis on reproductive organs. Dis. Aquat. Org. 30: 53-72. 11. Lo, C.F., Leu, J.H., Ho, C.H., Chen, C.H., Peng, S.E., Chen, Y.T., Chou, C.M., Yeh, P.Y., Huang, C.J., Chou, H.Y., Wang, C.H. & Kou, G.H. (1996) Detection of baculovirus associated with white spot syndrome (WSBV) in penaeid shrimp using polymerasechain reaction. Dis. Aquat. Org. 25: 133-141. 12. Lo, C.F., Ho, C.H., Peng, S.E., Chen, C.H., Hsu, H.C., Chiu, Y.L., Chang, C.F., liu, K.F., Su, M.S., Wang, C.H. & Kou, G.H. (1996) White spot syndrome baculovirus (WSBV) detection in cultured and captured shrimp, crabs and other arthropods. Dis. Aquat. Org. 27: 215-225. 13. Wang, C.H., Lo, C.F., Leu, J.H., Chou, C.M., Yeh, P.Y., Chou, H.Y., Tung, M.C., Chang, C.F., Su, M.S. & Kou, G.H. (1995) Purification and genomic analysis of baculovirus associated with white spot syndrome (WSBV) of Penaeus monodon. Dis. Aquat. Org. 23: 239-242. |
||
|
|
||
|
|
||
|
|
||
|
|
▲ TOP ▲
RBM3-8 ◢ 最近修訂日期︰ 2000/10/16