微生物學科 鄧 述 諄
| No. 3 (September, 2000) | ||
|
|
||
|
生物醫學報導 3 | |
| 染色體尾端之結構與重要性 | ||
|
微生物學科 鄧 述 諄 |
||
|
|
||
|
染色體尾端又稱端粒 (telomere),具有與染色體其他部分完全不同之結構與功能。這結構用以保護染色體的完整性及防止染色體重組 (recombination)。若是失去了這結構則染色體無法完全的複製 (replication)。在近五年來我們對於端粒複製機制的瞭解有長足的進步。這領域中所有的模型系統都被廣泛的研究。這幾年許多學術論文並證實端粒複製對於染色體穩定、老化以及癌症形成有重要的影響。 |
||
|
端粒的結構與端粒長度的控制 |
||
|
端粒通常是由短,但重複的序列所組成。這序列從 5' 端到 3' 端含高比率的 G 去氧核醣核酸。在酵母菌 Saccharomyces 的端粒便含有大約 350 個鹼基對 (base pair) 的 TG1-3去氧核醣核酸。而所有脊椎動物 (包括人類) 的端粒是由非常多重複的 TTAGGG 所組成。人類的染色體尾端便具有長達 5 kb 的 TTAGGG。 |
||
|
不完全的去氧核醣核酸複製可造成染色體減短 |
||
|
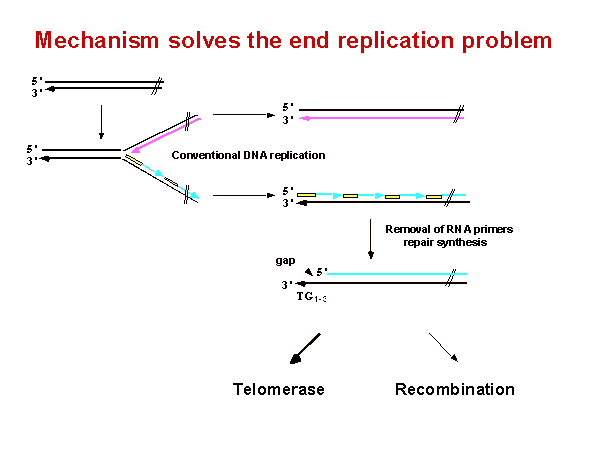
染色體尾端在複製過程中出現了一個問題 (見圖一)。在傳統的去氧核醣核酸複製中 leading strain 可以複製到染色體的最尾端。但是在 lagging strain 的複製時,由於起始的 8-12 bp 的 RNA 引子會被 RNase 吃掉,lagging strain 在每一次的染色體複製的過程中會損失 8-12 bp。倘若細胞缺乏一種增長端粒的能力,染色體便會越來越短,當短到進入 coding sequence 時便導致失去必須基因 (essential gene) 時,細胞便會死亡了。端粒通常具有特殊的蛋白質複合體 (protein complex) 保護著。當失去這些蛋白質複合體保護時,端粒便會出現不規則的長度。舉例來說,端粒複製機制我們對它的瞭解大多從酵母菌 (Saccharomyce) 這個模型系統來的。在酵母菌端粒含有 Rap1 蛋白質坐落在上。在一個受損的 Rap1 的細胞中,端粒由正常的 350 bp 增長到大約 500 bp。而在人類染色體端粒由 TRF1 及 TRF2 蛋白質複合體的保護,當 TRF2 缺乏時,大量的染色體融合便出現在染色體尾端。所以這些保護端粒的蛋白質具有穩定染色體的能力,並由他們來控制端粒的長度。 |
||
|
|
||
|
圖一 Mechanism solves the end replication problem |
||
|
端粒酵素為主要端粒複製時所使用的蛋白質 |
||
|
端粒大都為端粒酵素所生成。大部分的模型系統 (包括酵母菌及人類)的端粒酵素皆在近五年內 clone 到的。端粒酵素是由一個 RNA 及一個蛋白質所組成。這些 RNA 是端粒複製時的模板 (template);而蛋白質是一個反轉複製酵素 (reverse transcriptase)。他們利用染色體尾端的 3'-hydroxyl 當作引子 (primer) 而 RNA 當作模板,以反轉複製酵素去延長其端粒。舉個例子來說,人類端粒酵素的 RNA 含有一段 CCCTAA 序列在模板的中央。當反轉錄酵素用這模板生成新的端粒時, 便生成 TTAGGG 了。而每完成一次複製,反轉複製酵素便會面對一個能量阻礙 (energy barrier) 而無法繼續複製。這時反轉錄複製酵素需要經過一次轉位 (translocation) 而使得它的活性部位中心 (active site) 接近新生成的 3'-hydroxyl 引子,使得下一循環的端粒複製可以形成。所以依此方式可使得端粒增長。 |
||
|
端粒酵素缺乏時端粒重組為另一形成新端粒的能力 |
||
|
酵母菌以及其他真核生物一搬,當端粒酵素缺乏時,酵母菌的端粒便慢慢的變短,染色體失去,導致最後細胞死亡。然而,一部份的細胞即使在缺乏端粒酵素的狀況下仍可存活,這類細胞即為 DNA 重組能力正常的細胞。存活下來的酵母菌可經由去氧核醣核酸重組去增長端粒或端粒旁邊的序列,而形成長而不規則的端粒。 |
||
|
癌症細胞需要有無限制複製染色體的能力 |
||
|
在大部分人類的體細胞內並沒有端粒酵素的表達,所以他們的端粒隨著年紀的增長而逐漸變短。這種表現型非常類似端粒酵素缺乏的酵母菌,所以人類的體細胞都具有固定而有限的 passage;相反的,大部分的人類腫瘤細胞及細胞系可偵測到端粒酵素的表達,他們利用此酵素去維持端粒而使得染色體可以無限制的複製。然而一部份的腫瘤細胞和細胞系並無法偵測到端粒酵素的表達,卻類似端粒酵素缺乏而存活的酵母菌具有長而不規則的端粒。這些現象顯示可經由去氧核醣核酸重組去增長其端粒。而癌症細胞便運用端粒酵素或端粒重組去增長其端粒,而使其具有無限制複製染色體的能力。去瞭解端粒酵素正常和缺乏的酵母菌如何複製端粒將有助於了解端粒酵素具有及缺乏的腫瘤細胞與細胞系如何複製端粒。 |
||
|
抗端粒複製被視為新的癌症治療途徑 |
||
|
由於端粒複製可經由端粒酵素及端粒重組而達成端粒增長的部份,並且端粒複製與癌症細胞的無限制的複製有密切的關連,世界各國從學術界到製藥界已投入大量的金錢及人力去了解端粒複製的反應機制及可能的 drug target 用以發展抗癌的新藥。美國的 Geron 生技公司便於五年前置下一個酵母菌端粒酵素的抑制蛋白 Piflp 的專利,希望能進一步的由此開發新藥。美國的 Amgen 及 Geron 公司也利用反轉錄酵素的抑制藥去測試人類端粒酵素的活性,這個領域目前看似欣欣向榮,但也是問題重重。大部分的端粒酵素的調控因子 (regulator) 都尚未找尋 (clone) 到,只有酵母菌有三個端粒酵素的調控因子被找到。而他們對等的 (conserved) 人類蛋白目前仍為被尋找到。完全了解端粒酵素的反應機制仍來日方長。此外,Amgen 生技公司已發現一旦使用端粒酵素的抑制物於細胞上,細胞可由端粒重組可達成端粒增長的目的。所以目前科學界 (包括我個人的實驗室) 也致力於瞭解端粒重組的反應機制。希望不久的未來我們對這兩個端粒複製的反應機制有更完整的瞭解,進而發展出藥物可限制癌細胞端粒的生長,而使其失去無限制染色體複製的能力。 |
||
|
參考文獻 |
||
|
Biessmann, H., and Mason, J.M. (1997) Telomere maintenance without telomerase. Chromosoma 106, 63-69. Bryan, T.M., Anglezou, A., Gupta, J., Bacchetti, S., and Reddel, R.R. (1995) Telomere elongation in immortal human cells without detectable telomerase activity. EMBO J. 14, 4340-4348. Bryan, T.M., Englezou, A., Dalla-Pozza, L., Dunham, M.A., and Reddel, R.R. (1997) Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat. Med. 3, 1271-1274. Counter, C.M., Meyerson, M., Eaton, E.N., and Weinberg, R.A. (1997) The catalytic subunit of yeast telomerase. Proc. Natl. Acad. Sci. USA 94, 9202-9207. Hanahan, D., and Weinberg, R.A. (2000) The hallmarks of cancer. Cell 100, 57-70. Lendvay, T.S., Morris, D.K., Sah, J., Balasubramanian, B., and Lundblad, V. (1996) Senescence mutants of Saccharomyces cerevisiae with a defect in telomere replication identify three additional EST genes. Genetics 144, 1399-1412. Lingner, J., Cech, T.R., Hughes, T.R., and Lundblad, V. (1997) Three Ever Shorter Telomere (EST) genes are dispensable for in vitro yeast telomerase activity. Proc. Natl. Acad. Sci. USA 94, 11190-11195. Lingner, J., Hughes, T.R., Shevchenko, A., Mann, M., Lundblad, V., and Cech, T.R. (1997) Reverse transcriptase motifs in the catalytic subunit of telomerase. Science 276, 561-567. Lundblad, V., and Blackburn, E.H. (1993) An alternative pathway for yeast telomere maintenance rescues est- senescence. Cell 73, 347-360. Nakamura, T.M., Cooper, J.P., and Cech, T.R. (1998) Two modes of survival of fission yeast without telomerase. Science 282, 493-496. Nakamura, T.M., Morin, G.B., Chapman, K.B., Weinrich, S.L., Andrews, W.H., Lingner, J., Harley, C.B., and Cech, T.R. (1997) Telomerase catalytic subunit homologs from fission yeast and human. Science 277, 955-959. Nugent, C.I., Hughes, T.R., Lue, N.F., and Lundblad, V. (1996) Cdc13p: a single-strand telomeric DNA-binding protein with a dual role in yeast telomere maintenance. Science 274, 249-52. Singer, M.S., and Gottschling, D.E. (1994) TLC1, the template RNA component of the Saccharomyces cerevisiae telomerase. Science 266, 404-409. Teng, S.-C., and Zakian, V. (1999) Telomere-telomere recombination is an efficient bypass pathway for telomere maintenance in Saccharomyces cerevisiae. Mol. Cell. Biol. 19, 8083-8093. Zakian, V.A. (1995) Telomeres: beginning to understand the end. Science 270, 1601-1607. |
||
|
|
||
|
|
||
|
|
||
|
|
▲ TOP ▲
RBM3-6 ◢ 最近修訂日期︰ 2000/09/30